現在,マスメディアの取り扱いから,脳によい食品に対する注目度が高まっていることがうかがい知れる.しかし,脳と栄養素・食品の関係性の情報に関しては,経験則,また,科学的根拠の少ないものも多いのが実態である.一方,うつ病,統合失調症,心的外傷後ストレス障害といった精神疾患は,遺伝要因(遺伝子変異が原因)のみを原因とせず,環境要因との複合的な要因によって発症する.重要なことに,生活習慣病のみならず,これら精神疾患においても,食生活が重大な環境要因となっている.このような背景から,現在,脳機能と栄養素・食品との関連性が明確にされる必要に迫られており,さらに一歩進めて,栄養素・食品による脳機能の向上や疾患の治療・改善に向けた研究の需要も高まっているといえよう.
脳機能に対するその特異的役割,さらに,その作用機構が明確にされた必須栄養素はいまだ少ない.この理由として,古くから栄養素の研究分野で用いられてきた栄養素欠乏動物においては,欠乏動物自体が衰弱してしまい,脳機能異常を反映するような行動異常を検出しにくかった点があげられる.特に,病的な欠乏動物を用いては,正常な運動能力を必要とする学習・記憶能力や情動行動を測定することが困難である.また,欠乏動物を解析するための神経科学的手法や分子生物学的手法を中心とする解析方法が十分に整備されていなかった点もあげられる.しかし,現在,栄養素の標的遺伝子群等の同定が進み,これら遺伝子を対象とした脳特異的な遺伝子操作も可能であり,このような手法を用いて脳において擬似的栄養素欠乏状態を誘導することもできる.一方,1990年代以降,げっ歯類を個体レベルで解析する神経科学的手法が日進月歩に進み,イメージング,電気生理学,分子生物学,行動学などの多彩な手法を組み合わせて脳機能制御のメカニズムを統合的に理解するストラテジーが確立されてきた.以上のような,分子遺伝学的手法や神経科学的手法に起こったパラダイムシフトにより,現在,脳機能に対する栄養素および食品の役割や影響,また,その作用機構を研究する土台ができあがっている.たとえば,さまざまな栄養条件で飼育したマウスやラットの解析に,遺伝子操作マウスの解析用に確立されてきた手法をそのまま活用することが可能である.しかも,この方法を用いれば,遺伝子操作の影響と比較して,栄養素・食品の機能をより定量的に評価することもできる.
我々は,さまざまな神経科学的手法を活用して脳機能に対する必須栄養素群の役割の解明に取り組んできた.本稿では,ビタミンA,ビタミンB1,マグネシウム,必須アミノ酸トリプトファンなどの脳機能に対する役割を解析した研究,栄養不良による脳機能低下の共通メカニズム,さらに,今後の脳栄養学的研究の展望などを紹介する.
ビタミンB1(B1)は鈴木梅太郎博士が約100年前に最初に発見したビタミンである.B1は糖代謝系やATP産生に対する補酵素として機能し,グルコースがエネルギー源となる脳では,ATP産生に対するB1の重要性が特に高い.一方,B1欠乏症状として脚気が有名であるように,B1は神経系の機能維持にも必要不可欠である.特に重要な点として,B1欠乏による重篤な疾患であるウェルニッケ・コルサコフ症候群が知られており,この疾患の顕著な病態は記憶障害である.脚気はB1摂取により速やかに改善されるものの,ウェルニッケ・コルサコフ症候群における記憶障害は不可逆的であり,B1欠乏の後遺症となっている.B1欠乏により記憶障害が導かれることは古くから知られていたが,この記憶障害が引き起こされるメカニズムは不明であった.
現在,脚気という言葉はほとんど聞かれなくなっている.しかし,B1欠乏によるウェルニッケ・コルサコフ症候群はみられなくなったわけではない.国内のアルコール依存症患者は100万人以上であり,この中の少なくとも数%には記憶障害が観察されるが,この記憶障害に至る原因がB1欠乏であるといわれている1, 2).一方,アルツハイマー型認知症発症過程にもB1欠乏が関連すると唱えられている2, 3).以上のように,アルコール依存症における記憶障害は「隠れB1欠乏症」であり,B1欠乏による記憶障害は意外にも身近な疾患といえよう.
我々は,B1欠乏による記憶障害のメカニズムの解明を進めてきた4).野生型マウス(C57BL/6N系統)にB1欠乏飼料を10日間給餌し,この間ビタミンB1拮抗物質であるピリチアミンを連日投与すると,マウスは病的な状態に陥るものの,この後通常食を3週間摂取させると,対照群と同程度まで運動協調性や行動活性が回復した.しかし,このB1欠乏から回復させたマウス(B1欠乏回復マウス)では,モリス水迷路による空間記憶,他のマウスを記憶する社会記憶,また,恐怖条件づけ文脈記憶などの海馬依存性記憶形成能力の顕著な障害が認められた(表1).この記憶障害はB1欠乏から回復後も少なくとも6か月以上持続し,ウェルニッケ・コルサコフ症候群と同様に,マウスにおいてもB1欠乏は不可逆的な記憶障害を誘導することが示された.一方,B1欠乏回復マウスの扁桃体依存的記憶形成能力は正常であり,B1欠乏は海馬依存性記憶の障害を導くことが示唆された.重要なことに,B1欠乏経験後の記憶障害は,我々がこれまでに解析を進めてきた記憶に重要な分子群の阻害や遺伝子欠損に匹敵する程度のものであった.以上の研究結果から,B1欠乏回復マウスは慢性的な記憶障害を示すウェルニッケ・コルサコフ症候群モデルであること,また,この記憶障害は海馬の機能低下と関連することが強く示唆された.
表1 栄養素摂取不足による記憶障害と神経変性 | 海馬依存性記憶 | 扁桃体依存性記憶 | 脳内炎症 | 海馬変性 | CREB情報伝達活性 |
|---|
| サイトカイン発現量 | ミクログリア活性 | 細胞数 | スパイン数 |
|---|
| B1欠乏 | 障害 | 正常 |  |  |  |  |  |
| Mg欠乏 | 障害 | 正常 |  |  |  |  |  |
| TRP制限 | 一部障害 | 正常 | N.D. | N.D. | N.D. | N.D. | N.D. |
| 高脂肪食 | 障害 | N.D. | N.D. |  | N.D. |  | N.D. |
| B1欠乏,Mg欠乏,TRP制限の結果は我々のグループの結果をまとめたものである(本文参照).高脂肪食は他のグループの結果である22).N.D.: not determined. |
海馬はエピソード記憶形成の中枢であり,学習・記憶をつかさどる中心的な領野である.そこで,B1欠乏が海馬に与えるダメージを形態学的に解析した.その結果,B1欠乏回復マウス海馬では顕著な神経細胞数の低下が観察され,B1欠乏により海馬ニューロンが欠落することが示唆された.続いて,神経細胞特異的にGFPを発現するThy1-GFPマウスにB1欠乏処置を施して,海馬ニューロンの形態を解析した結果,B1欠乏回復マウスの海馬歯状回では,ニューロンの樹状突起スパイン密度の顕著な減少が観察された.以上より,B1欠乏により,海馬ニューロンの変性,すなわち,海馬の神経変性が引き起こされ,その結果,海馬依存的記憶の障害が導かれることが強く示唆された(表1)4).
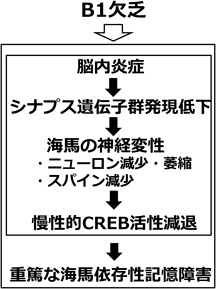
現在,B1欠乏が海馬の変性を導く分子メカニズムの解析を進めている.未発表であるため,詳細は省くが,次世代シークエンサーを用いたRNA-Seq解析から,B1欠乏時の海馬ではまず炎症性サイトカイン群の高発現が観察され,引き続き,シナプス関連遺伝子群の発現も低下することが明らかとなった.さらに,B1欠乏回復以降も,記憶形成に必須な遺伝子発現を制御する転写調節因子CREB(cAMP response element binding protein)情報伝達経路に障害が残ることも示唆された.以上の結果から,B1欠乏により海馬において強い脳内炎症が誘導され,その後,炎症による海馬組織の損傷が誘導され,最終的にCREBの活性が損なわれ,記憶障害に至る機構が示唆されている.言い換えれば,海馬はエネルギー消費の高い脳領野であるため,B1欠乏によるダメージを受けやすく,B1欠乏状態に陥ると劇的な脳内炎症が起こり,海馬が変性し,その結果として,記憶障害が引き起こされるものと考えられる(図1).以上のように,長い間不明であったB1欠乏による記憶障害のメカニズムの解明までもう一歩のところまで近づいている.
ビタミンA(レチノール)は生体内でall-transレチノイン酸(all-trans retinoic acid:ATRA)とこの異性体である9-cisレチノイン酸(9-cis RA)に代謝される.ATRAと9-cisRAは核内受容体であるレチノイン酸受容体(retinoic acid receptor:RAR)とレチノイドX受容体(retinoid X receptor:RXR)のリガンドであり,これら受容体による標的遺伝子群の転写制御を介して生理作用を発揮する.RARとRXRにはそれぞれにα, β, γの3種類のサブタイプが存在し,ATRAはRARの,一方,9-cis RAはRARとRXRのリガンドとなる.
ビタミンA欠乏症状として夜盲症が有名である.一方,レチノイン酸は神経系,特に脊髄の発生・分化に重要な役割を果たしている.これらの点はビタミンAと脳神経系との密接な関係を示唆するものである.成体の脳においても,RARとRXRのほとんどのサブタイプの発現が認められ,すべてのサブタイプの発現を合わせると脳のほとんどの領域でRARおよびRXRが高発現している.したがって,ビタミンA−レチノイン酸情報伝達経路が脳機能に対しても重要な役割を担っていることが推察される.しかし,脳高次機能に対するビタミンAの役割は栄養学的には明らかにされてこなかった.この点は,冒頭に記したように,ビタミンA欠乏により動物は病的な状態に陥るため,脳機能が変化していたとしても,観察しにくかったためではないかと考えられる.しかし,RARおよびRXRなどの遺伝子改変マウス群の解析が進み,脳高次機能に対するビタミンAの重要性が明らかになりつつある5).
他のグループの解析から,RARβ遺伝子欠損マウスとRARβ/RXRγ二重遺伝子欠損マウスを用いて,細胞レベルにおける学習・記憶のモデルと捉えられている長期増強(long-term potentiation:LTP)および長期抑圧(long-term depression:LTD)が解析された結果,これら遺伝子欠損マウスの海馬CA1領域では,シナプス伝達(basal synaptic transmission)は正常であるものの,LTPおよびLTDがほとんど観察されなくなることが明らかになった6).この結果に一致して,ビタミンA欠乏マウスの海馬CA1領域においてもLTPとLTDに障害が観察されることが示唆された7).以上の結果から,レチノイン酸受容体の遺伝子欠損マウスやビタミンA欠乏マウスの海馬の興奮性ニューロンでは,記憶形成を導くような強い刺激がもたらされても神経可塑的な変化は起こらない,すなわち,記憶を形成できないことが示唆された.
先述したRARβ遺伝子欠損マウスとRARβ/RXRγ二重遺伝子欠損マウスには顕著な運動能力の低下が観察されたため,正常な運動能力を必要とする学習記憶課題を用いて,これら遺伝子欠損マウスの記憶能力を正当に評価することができなかった.そこで,我々は,学習・記憶に対するビタミンA情報伝達経路の役割を明らかにするために,海馬を含む前脳領域特異的なRARのコンディショナル変異マウスを作製することで,脳部位特異的に擬似的なビタミンA欠乏状態を誘導して,その影響を解析した(表2)8).時期特異的な遺伝子発現誘導を可能にするテトラサイクリンシステムを利用して,ドミナントネガティブ型RARα変異体(dnRAR)を前脳領域特異的に発現誘導させるトランスジェニックマウスを作製し,成体になってから前脳領域にdnRARを発現させてレチノイン酸受容体群による転写制御を阻害した影響を解析した.重要な点として,この変異マウスでは,モリス水迷路における空間記憶,初対面のマウスを記憶する社会記憶などの海馬依存性記憶形成に障害が観察された.一方,レチノイン酸受容体群の全身性の遺伝子欠損マウスの結果に一致して,このdnRAR発現マウスにおいても海馬CA1領域のLTPに障害が観察された.興味深い点として,dnRAR発現マウスでは1回のテタヌス刺激では海馬LTPに障害が観察されたものの,4回テタヌス刺激を与えることで,この障害は解消され,野生型マウスと同程度のLTPが観察された.これに対して,dnRARマウスは,社会記憶課題において,初対面のマウスを一度だけ3分間対面させた場合には,このマウスを24時間後には覚えていなかったのに対して,1時間間隔で二度,3分間ずつ対面させた場合(分散学習“spaced training”と呼ばれている学習手法)には,野生型マウスと同様に24時間後にも相手のマウスを覚えていた.このように,強い刺激を与えることで海馬LTPと海馬依存性社会記憶の障害が解消されたことから,dnRARマウスでは海馬神経可塑的変化と海馬依存性記憶が相関的な関係であることが強く示唆され,ビタミンA情報伝達経路は海馬神経可塑性を制御することで記憶を制御すると考えられた(表2).また,レチノイン酸受容体のアンタゴニストを海馬に注入しても同様に,社会記憶に障害が観察された.以上までの結果から,ビタミンAが海馬における記憶制御に必須な役割を果たしていることが示唆された8).
表2 前脳特異的ビタミンA情報伝達経路改変マウスの解析結果のまとめ| ビタミンA情報伝達 | 発現受容体(RAR) | 電気生理学的解析 | 海馬依存性記憶課題 |
|---|
| 基本的シナプス伝達(Basal Synaptic Transmission) | 長期増強(LTP) | 空間記憶(モリス水迷路課題) | 恐怖記憶(恐怖条件づけ文脈記憶) | 社会記憶(社会的認知記憶課題) |
|---|
| 100 Hz |
|---|
| ×1 | ×4 | | 分散学習 |
|---|
| 不活性化 | dnRAR | 正常 | 障害 | 障害改善 | 障害 | 障害 | 障害 | 障害改善 |
| 活性化 | RARα | 正常 | N.D. | N.D. | 向上 | N.D. | 向上 | N.D. |
| ドミナントネガティブRAR変異体あるいは野生型RARαを前脳領域特異的に発現したトランスジェニックマウスの解析結果をまとめた(本文参照).dnRAR:dominant negative mutant RAR, N.D:not determined. |
一方,レチノイン酸受容体のアゴニストを海馬に注入した場合,社会記憶能力の向上が観察された.実際には,初対面のマウスに1.5分間対面させた場合には,対照群のマウスは24時間後にこのマウスを覚えていなかったが,アゴニストを注入されたマウスは24時間経過しても相手のことを覚えていた(未発表データ).さらに,現在,RARαを前脳領域特異的に過剰発現させたマウスを作製し,その解析を進めているが,このトランスジェニックマウスにおいても記憶能力の向上が観察されている(未発表データ).以上の解析から,ビタミンA情報伝達経路の“loss of function”により記憶能力が低下し,一方,“gain of function ”により記憶能力が向上することが明らかになりつつある(表2).したがって,ビタミンAは記憶能力を正に制御する栄養素であるという結論がみえてきている.現在,次世代シークエンサーを用いたRNA-Seq解析によりレチノイン酸受容体群の標的遺伝子群の同定を進めており,候補遺伝子が見つかっている.今後,この遺伝子群の機能解析を進めることで,ビタミンAによる記憶制御の分子機構の解析を進める予定である.
他方,他のグループの報告より,加齢に伴う記憶能力低下とビタミンA情報伝達経路の活性低下が相関を示すことが示唆されている.加齢に伴い脳内RARβおよびRXRβ/γ mRNA量が低下することから,加齢によるビタミンA情報伝達の減退が示唆され,一方,老齢マウスにビタミンAやATRAを投与すると,加齢に伴う記憶能力低下と海馬CA1領域のLTPの低下などが阻止される9, 10).したがって,ビタミンA情報伝達経路の減弱が老化に伴う記憶能力の低下の一因となる可能性がある.さらに,アルツハイマー型認知症モデルマウスにATRAを投与することで,この疾患の病理学的マーカーであるアミロイドβ(amyloid β:Aβ)の沈着が抑制され,記憶障害も改善されることが示唆されている11).
RARβ/RXRβあるいはRARβ/RXRγの二重遺伝子欠損マウスでは,行動活性の低下とドーパミン受容体D2R(D2R)の発現量の低下が認められた12).また,RXRγあるいはRXRβ/RXRγ遺伝子欠損マウスでは強制水泳試験およびスクロース嗜好性試験においてうつ病様行動が観察された13).これら遺伝子欠損マウスの側坐核シェル領域ではD2Rの発現量の低下が観察され,この領域におけるRXRγあるいはドーパミン受容体の発現の回復によりうつ病様行動が解消された13).以上の結果は,側坐核シェル領域におけるビタミンA情報伝達経路がD2R遺伝子の転写活性化を介して情動・気分行動を制御することを示唆している.RARおよびRXRのサブタイプは脳内のさまざまな領野において特異的な発現を示すため,ビタミンAはそれぞれの脳領野特異的な機能の制御に関わることが予想され,結果的に広範囲の脳高次機能に関わるものと考えられる.
我々のグループの研究を含めた以上までの知見は,ビタミンAが記憶能力を正に制御することを示しており,分子レベルの解析による制御機構の実態解明が待たれる.さらに,ビタミンAが記憶増強因子(memory enhancer)として,加齢による記憶能力低下やアルツハイマー型認知症の治療・防止に有効である可能性も大きい.ビタミンAの過剰症に対する対策は必要であるものの,ビタミンAの有用性は高そうであり,疾患対策など応用面での活用の検討が必要である.
脳では,アミノ酸は神経伝達物質およびその前駆体として脳高次機能制御に大きな役割を担っている.この点は他の組織におけるアミノ酸の役割と一線を画する点である.
グルタミン酸は神経伝達物質として機能し,グルタミン酸受容体に結合して,記憶を中心とするさまざまな脳機能を制御する.重要なことに,必須アミノ酸であるトリプトファン,ヒスチジンはそれぞれ神経伝達物質セロトニンとヒスタミンの前駆体であり,これらの神経伝達物質の脳内量は摂食量の影響を直接的に受けている.
セロトニンは怒り,攻撃性や不安などの情動行動を中心として,学習・記憶,食欲,睡眠など広範な脳機能制御に関わる.トリプトファンはトリプトファン水酸化酵素と芳香族L-アミノ酸脱炭素酵素による2段階の酵素反応により5-ヒドロキシトリプトファンを経てセロトニンへと代謝される.脳内トリプトファン濃度は代謝酵素の反応を飽和させるまでに至っていないため,トリプトファン摂取量,脳内トリプトファン濃度とセロトニン量は正の相関を示す14–16).したがって,トリプトファン摂取量は脳内セロトニン量を決定しており,トリプトファンはセロトニンの作用に間接的に影響を及ぼしている.我々は,トリプトファン摂取不足が脳高次機能に与える影響を明らかにするため,マウスにトリプトファン量を35%減らしたマイルドな制限食(低トリプトファン食)を給餌し,その影響を解析した17, 18).この条件では,摂取制限を半年続けてもマウスは外見上健康であり,体重異常も観察されない.行動バッテリー解析を用いて,行動レベルの変化を解析したところ,トリプトファン摂取制限により,社会記憶や空間記憶には異常が観察されなかったものの,海馬依存性の恐怖条件付け文脈記憶に異常が観察された(表1)17).一方,情動行動に関しては,オープンフィールド試験と強制水泳試験において行動活性の増加が観察され,新規環境や水中では躁様の行動を示したことから,トリプトファン摂取制限により情動行動にも異常が現れることが明らかとなった18).
興味深いことに,トリプトファン摂取制限により,相手のマウスに対して先制攻撃を加える攻撃性(offensive aggression)に異常はみられなかったものの,先制攻撃を受けた後に反撃する攻撃性(defensive aggression)が強くなっていた18).セロトニン量の低下により攻撃性が増加することが知られているため,トリプトファン摂取不足によるセロトニン量の低下により攻撃性が増加したことが示唆された.さらに,上述のように,トリプトファン摂取制限により恐怖反応が低下しており,また,躁様になっているため,先制攻撃を受けてもひるむことなく他者を攻撃してしまう「無鉄砲」な状態になっている可能性が考えられた.
以上のように,トリプトファン摂取制限による情動行動異常が観察されたものの,ビタミンAやB1の解析で観察されたような強い行動変化は観察されなかった.このことは摂取制限(35%減)の条件によるものであると考えられ,決してトリプトファンの役割が小さいことを示すものではない.今後,トリプトファンとそれ以外のアミノ酸の役割も含めて,手法や視点を変えて解析することで興味深い結果が得られてくるように思われる.
ミネラルはイオンチャネルや酵素の活性調節に必要であるため,脳機能にも大きな影響を及ぼすことが予想される.マグネシウムは300以上の酵素のコファクターとして働き,さまざまな組織において必要不可欠なミネラルである.
NMDA型グルタミン酸受容体は,神経伝達物質グルタミン酸の受容体であり,リガンドかつ電位依存性カルシウムチャネルとして,記憶を形成させる強い刺激に応答して,細胞内カルシウム情報伝達経路を活性化する.マグネシウムイオンはNMDA型グルタミン酸受容体の“Mg2+ブロック”を担っており,脱分極に反応して受容体から外れて,細胞外からのカルシウムイオン流入を可能にする.脳内のマグネシウムイオン濃度の上昇は学習・記憶能力を高めることが示唆されており19),一方,マグネシウム欠乏は記憶障害を導くことが示唆されている20).
我々はマウスを用いて食餌性マグネシウム欠乏の影響を,ビタミンB1欠乏マウスやトリプトファン摂取制限マウスと同様の行動バッテリーにより解析した(表2)21).食餌性マグネシウム欠乏を5週間経験したマウスでは,扁桃体依存性記憶や行動活性に異常は観察されなかったものの,モリス水迷路における空間記憶,恐怖条件づけ文脈記憶,社会記憶の海馬依存性記憶に障害が観察された.しかし,海馬,扁桃体,前頭前野のニューロン樹状突起の解析を行ったが,スパイン密度とスパインの形態には異常は観察されなかった.したがって,マグネシウム欠乏は,B1欠乏と同様に海馬依存性記憶の障害を導くものの,B1欠乏とは異なり海馬ニューロンの形態には影響を及ぼさないことが示された.
さらに,マグネシウム欠乏による記憶障害のメカニズムを理解するために,次世代シークエンサーを用いたRNA-Seq解析を行ったところ,Gene Ontology解析からマグネシム欠乏により炎症関連遺伝子群の発現が増加することが明らかとなった.定量的RT-PCRを用いた解析からも,炎症性サイトカイン等の顕著な発現が観察され,マグネシウム欠乏によって脳内炎症が起こることが強く示唆された.しかしながら,ビタミンB1欠乏の場合に比較すると,マグネシウム欠乏ではB1欠乏時ほど,炎症関連遺伝子群の発現は劇的に増加しておらず,軽度の脳内炎症が起こったと考察している(表2,投稿準備中).
興味深いことに,他のグループの解析から,高脂肪の食餌を摂取させたマウスにおいても,海馬依存性記憶障害,海馬ニューロンの形態異常,強い脳内炎症が観察されている22).そこで,これまでの知見を総合すると,B1欠乏や高脂肪食摂取の場合のように脳内炎症が強くなるとニューロンの欠落や形態変化など組織の損傷が大きくなるのではないかと考えられる.また,栄養状態の悪化により共通して脳内炎症が誘導される可能性が大きく,栄養状態の悪化と脳内炎症の相関性を解析することも重要であると考えている.
近年,腸内細菌に注目が集まっており,脳の記憶機能に対してもその重要性が示唆されている23).我々は,マウスに乳酸菌の死菌を1か月食餌として与えることでその影響を解析した24).その結果,乳酸菌を摂取したマウスは海馬依存性の社会記憶の向上を示した.一方,海馬歯状回では成体になっても神経細胞の産生が行われているが,この神経新生も乳酸菌摂取により促進されることが明らかとなった.我々の過去の解析では,NMDA型グルタミン酸受容体のアンタゴニストであるメマンチン投与により人為的に神経新生を促進させた場合にも海馬依存性記憶能力の向上が観察されている25).そこで,乳酸菌摂取により,海馬神経新生の促進を通して記憶能力が向上した可能性が考えられ,腸内細菌が記憶増強因子となる可能性が示された.
神経新生により産生された若いニューロンが記憶能力向上を導く一方で,海馬神経新生の促進は既存の記憶回路を破壊することで,記憶の忘却を導くことが明らかにされている26, 27).そこで,この原理を利用して,トラウマ記憶の忘却を誘導することで,トラウマ記憶を原因とする心的外傷後ストレス障害(posttraumatic stress disorder:PTSD)の治療方法を開発することを試みている26–28).このような乳酸菌摂取による海馬神経新生の促進効果をPTSDの治療方法開発にも利用できる可能性が考えられる.
現在までの研究から,ビタミンA,ビタミンB1,トリプトファン,マグネシウムなどの摂取不足により記憶機能が障害を受けることが明らかとなりつつある.この中で,我々や他のグループの解析からビタミンAは記憶能力を向上させることが示唆され,栄養素の中でもビタミンAは別格であり,記憶制御プロセスを直接的に調節することが示唆されている.では「なぜ栄養素摂取不足の影響が記憶に集中するのか?」との印象を受けるかもしれない.記憶は我々の最も重要かつ高次な認知機能の一つであり,五感で感じ取った感覚から,心の動き(情動)に至るまでをすべて統合して記録する役割を果たしている.そのため,一つのエピソードを記憶するために,脳内の多領野が関わり,多数のニューロン内の多くの素過程からなる多段階の反応を必要としている.そこで,ニューロン機能の小さなほころびであろうとも,記憶能力には大きな影響が及んでしまうと考えられよう.
冒頭に記したように,食物摂取は精神疾患や神経変性疾患に対する重要な環境要因である.たとえば,精神医学領域における有名な疫学調査の結果として,魚を食べる国ほど大うつ病の有病率が少ないことが示されている29).この調査が契機となり,子供の知能指数と妊娠中の母親のω3不飽和脂肪酸摂取量の関係性も指摘され,脳機能に対するω3不飽和脂肪酸の重要性が示唆されている30).しかし,この一連の研究は例外的であり,それぞれの疾患の発症や病態と食物摂取の因果関係は十分に明らかになっていない.これは単に栄養学的な研究が進展していないためではなく,精神・神経変性疾患のメカニズムにもいまだに謎が多いことも原因である.したがって,疾患群のメカニズムの解明が進み,このメカニズムにおける栄養素・食品が果たす役割や及ぼす影響が解明される必要がある.一方,脳機能に対するω3不飽和脂肪酸の重要性は指摘されているものの,ω3不飽和脂肪酸の作用メカニズムの解明は進んでいない.そこで,上述の疫学調査から得られた大きな成果に応えるためにも,このメカニズムを明らかにする基礎研究の進展も必須である.以上のような方向性の研究が進めば,疾患のメカニズムに基づいた栄養素・食品の有効な活用の道が開けると考えられる.
今から20年ほど前,ビタミンB1欠乏によって海馬依存性記憶が障害を受けることを最初にいい始めたころ,「B1欠乏に海馬が関係するとは聞いたことがない」といわれ,かなりの塩対応を受けたことを記憶している.そのころ国内では「海馬依存性記憶課題」も知られておらず,神経科学的な手法も普及していなかった.そのときに比べると,この間の脳科学の進歩は目覚ましい.20年前には夢のようであった解析を現在はいとも簡単に行える.今後,脳と栄養素・食品との関係性を解明する脳栄養学的な研究が飛躍的に進展することを期待したい.
引用文献References
1) Akhouri, S., Kuhn, J., & Newton, E.J.(2020) in StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing.
2) Venkataraman, A., Kalk, N., Sewell, G., Ritchie, C.W., & Lingford-Hughes, A. (2017) Alcohol and Alzheimer’s Disease-Does Alcohol Dependence Contribute to Beta-Amyloid Deposition, Neuroinflammation and Neurodegeneration in Alzheimer’s Disease? Alcohol Alcohol., 52, 151–158.
3) Gibson, G.E., Hirsch, J.A., Fonzetti, P., Jordon, B.D., Cirio, R.T., & Elder, J. (2016) Vitamin B1 (thiamine) and dementia. Ann. N. Y. Acad. Sci., 1367, 21–30.
4) Inaba, H., Kishimoto, T., Oishi, S., Nagata, K., Hasegawa, S., Watanabe, T., & Kida, S. (2016) Vitamin B1-deficient mice show impairment of hippocampus-dependent memory formation and loss of hippocampal neurons and dendritic spines: potential microendophenotypes of Wernicke-Korsakoff syndrome. Biosci. Biotechnol. Biochem., 80, 2425–2436.
5) Maden, M. (2007) Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat. Rev. Neurosci., 8, 755–765.
6) Chiang, M.Y., Misner, D., Kempermann, G., Schikorski, T., Giguère, V., Sucov, H.M., Gage, F.H., Stevens, C.F., & Evans, R.M. (1998) An essential role for retinoid receptors RARbeta and RXRgamma in long-term potentiation and depression. Neuron, 21, 1353–1361.
7) Misner, D.L., Jacobs, S., Shimizu, Y., de Urquiza, A.M., Solomin, L., Perlmann, T., De Luca, L.M., Stevens, C.F., & Evans, R.M. (2001) Vitamin A deprivation results in reversible loss of hippocampal long-term synaptic plasticity. Proc. Natl. Acad. Sci. USA, 98, 11714–11719.
8) Nomoto, M., Takeda, Y., Uchida, S., Mitsuda, K., Enomoto, H., Saito, K., Choi, T., Watabe, A.M., Kobayashi, S., Masushige, S., et al. (2012) Dysfunction of the RAR/RXR signaling pathway in the forebrain impairs hippocampal memory and synaptic plasticity. Mol. Brain, 5, 8.
9) Etchamendy, N., Enderlin, V., Marighetto, A., Vouimba, R.M., Pallet, V., Jaffard, R., & Higueret, P. (2001) Alleviation of a selective age-related relational memory deficit in mice by pharmacologically induced normalization of brain retinoid signaling. J. Neurosci., 21, 6423–6429.
10) Mingaud, F., Mormede, C., Etchamendy, N., Mons, N., Niedergang, B., Wietrzych, M., Pallet, V., Jaffard, R., Krezel, W., Higueret, P., et al. (2008) Retinoid hyposignaling contributes to aging-related decline in hippocampal function in short-term/working memory organization and long-term declarative memory encoding in mice. J. Neurosci., 28, 279–291.
11) Ding, Y., Qiao, A., Wang, Z., Goodwin, J.S., Lee, E.S., Block, M.L., Allsbrook, M., McDonald, M.P., & Fan, G.H. (2008) Retinoic acid attenuates beta-amyloid deposition and rescues memory deficits in an Alzheimer’s disease transgenic mouse model. J. Neurosci., 28, 11622–11634.
12) Krezel, W., Ghyselinck, N., Samad, T.A., Dupé, V., Kastner, P., Borrelli, E., & Chambon, P. (1998) Impaired locomotion and dopamine signaling in retinoid receptor mutant mice. Science, 279, 863–867.
13) Krzyzosiak, A., Szyszka-Niagolov, M., Wietrzych, M., Gobaille, S., Muramatsu, S., & Krezel, W. (2010) Retinoid x receptor gamma control of affective behaviors involves dopaminergic signaling in mice. Neuron, 66, 908–920.
14) Gessa, G.L., Biggio, G., Fadda, F., Corsini, G.U., & Tagliamonte, A. (1974) Effect of the oral administration of tryptophan-free amino acid mixtures on serum tryptophan, brain tryptophan and serotonin metabolism. J. Neurochem., 22, 869–870.
15) Gessa, G.L., Biggio, G., Fadda, F., Corsini, G.U., & Tagliamonte, A. (1975) Tryptophan-free diet: a new means for rapidly decreasing brain tryptophan content and serotonin synthesis. Acta Vitaminol. Enzymol., 29, 72–78.
16) Stancampiano, R., Melis, F., Sarais, L., Cocco, S., Cugusi, C., & Fadda, F. (1997) Acute administration of a tryptophan-free amino acid mixture decreases 5-HT release in rat hippocampus in vivo. Am. J. Physiol., 272, R991–R994.
17) Uchida, S., Umeeda, H., Kitamoto, A., Masushige, S., & Kida, S. (2007) Chronic reduction in dietary tryptophan leads to a selective impairment of contextual fear memory in mice. Brain Res., 1149, 149–156.
18) Uchida, A., Kitamoto, H., Umeeda, N., Nakagawa, S., Masushige, S., & Kida, S. (2005) Chronic reduction in dietary tryptophan leads to changes in the emotional response to stress in mice. J. Nutr. Sci. Vitaminol. (Tokyo), 51, 175–181.
19) Slutsky, I., Abumaria, N., Wu, L.J., Huang, C., Zhang, L., Li, B., Zhao, X., Govindarajan, A., Zhao, M.G., Zhuo, M., et al. (2010) Enhancement of learning and memory by elevating brain magnesium. Neuron, 65, 165–177.
20) Bardgett, M.E., Schultheis, P.J., McGill, D.L., Richmond, R.E., & Wagge, J.R. (2005) Magnesium deficiency impairs fear conditioning in mice. Brain Res., 1038, 100–106.
21) Serita, T., Miyahara, M., Tanimizu, T., Takahashi, S., Oishi, S., Nagayoshi, T., Tsuji, R., Inoue, H., Uehara, M., & Kida, S. (2019) Dietary magnesium deficiency impairs hippocampus-dependent memories without changes in the spine density and morphology of hippocampal neurons in mice. Brain Res. Bull., 144, 149–157.
22) Cope, E.C., LaMarca, E.A., Monari, P.K., Olson, L.B., Martinez, S., Zych, A.D., Katchur, N.J., & Gould, E. (2018) Microglia Play an Active Role in Obesity-Associated Cognitive Decline. J. Neurosci., 38, 8889–8904.
23) Buffington, S.A., Di Prisco, G.V., Auchtung, T.A., Ajami, N.J., Petrosino, J.F., & Costa-Mattioli, M. (2016) Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell, 165, 1762–1775.
24) Ishikawa, R., Fukushima, H., Nakakita, Y., Kado, H., & Kida, S. (2019) Dietary heat-killed Lactobacillus brevis SBC8803 (SBL88™) improves hippocampus-dependent memory performance and adult hippocampal neurogenesis. Neuropsychopharmacol Rep, 39, 140–145.
25) Ishikawa, R., Kim, R., Namba, T., Kohsaka, S., Uchino, S., & Kida, S. (2014) Time-dependent enhancement of hippocampus-dependent memory after treatment with memantine: Implications for enhanced hippocampal adult neurogenesis. Hippocampus, 24, 784–793.
26) Ishikawa, R., Fukushima, H., Frankland, P.W., & Kida, S. (2016) Hippocampal neurogenesis enhancers promote forgetting of remote fear memory after hippocampal reactivation by retrieval. eLife, 5, e17464.
27) Ishikawa, R., Uchida, C., Kitaoka, S., Furuyashiki, T., & Kida, S. (2019) Improvement of PTSD-like behavior by the forgetting effect of hippocampal neurogenesis enhancer memantine in a social defeat stress paradigm. Mol. Brain, 12, 68.
28) Kida, S. (2019) Reconsolidation/Destabilization, Extinction and Forgetting of Fear Memory as Therapeutic Targets for PTSD. Psychopharmacology (Berl.), 236, 49–57.
29) Hibbeln, J.R. (1998) Fish consumption and major depression. Lancet, 351, 1213.
30) Hibbeln, J.R., Davis, J.M., Steer, C., Emmett, P., Rogers, I., Williams, C., & Golding, J. (2007) Maternal seafood consumption in pregnancy and neurodevelopmental outcomes in childhood (ALSPAC study): an observational cohort study. Lancet, 369, 578–585.
著者紹介Author Profile
 喜田 聡(きだ さとし)
喜田 聡(きだ さとし)東京大学大学院農学生命科学研究科応用生命化学専攻教授.博士(農学).
略歴1989年東京大学農学部卒業,94年同大学院農学研究科修了,日本学術振興会特別研究員,Cold Spring Harbor研究所ポスドクを経て,97~2019年東京農業大学(講師,02年助教授,08年教授).19年4月より現職.12~18年新学術領域「マイクロ精神病態」領域代表.
研究テーマと抱負記憶制御基盤の解明と脳疾患(PTSDや認知症など)の治療方法開発,記憶制御に対する栄養素の役割,食記憶による食認知行動制御基盤の解明を中心に認知機能のメカニズムを回路・細胞・分子レベルで研究している.
ウェブサイトhttps://kida-lab.org
趣味サッカー,散策,旅行,飲み会.