脂肪組織はエネルギーの貯蔵を主要な生理的役割の一つとしているため,当然のことながら食との関わりは大きい.食べ過ぎれば余剰なエネルギーは白色脂肪組織に蓄積され,肥満やメタボリックシンドロームを引き起こす.厚生労働省の国民健康・栄養調査によると,40~74歳では日本人男性の2人に1人が,女性の5人に1人がメタボリックシンドロームまたはその予備群である.内臓脂肪型肥満に加え,高血糖,高血圧,脂質異常症のうち二つ以上の症状を併せ持つメタボリックシンドームでは,心臓病や脳卒中などの動脈硬化性疾患をまねきやすく,その増加は大きな社会問題となっている.
メタボリックシンドロームの最大のリスクファクターである肥満症の対策が求められており,エネルギー消費の側面からの肥満対策のターゲットとして褐色脂肪が関心を集めている.褐色脂肪の活性化が肥満を軽減することは多くの動物実験により示されており,2019年に我々を含めた複数のグループから成人に機能的な褐色脂肪が存在することが示されたことにより1, 2),ヒトの肥満対策のターゲットとして関心が高まり急速に研究が進展している.
本稿でははじめに2種類の脂肪組織の機能と生理的役割について概説し,後半では食事や食品成分が脂肪組織に与える影響,そして脂肪組織が食に与える影響について紹介する.
1)白色脂肪組織と褐色脂肪組織
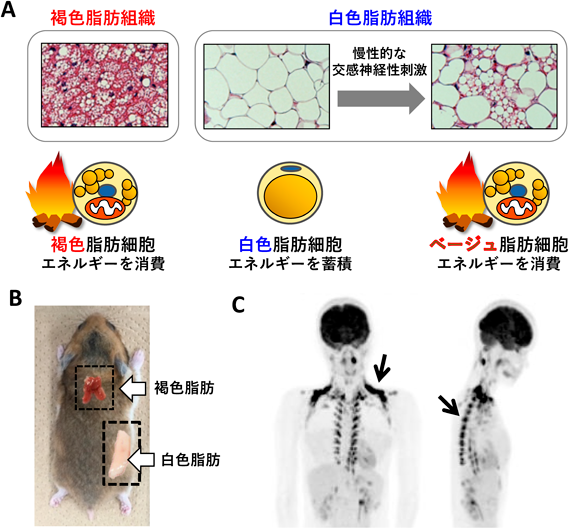
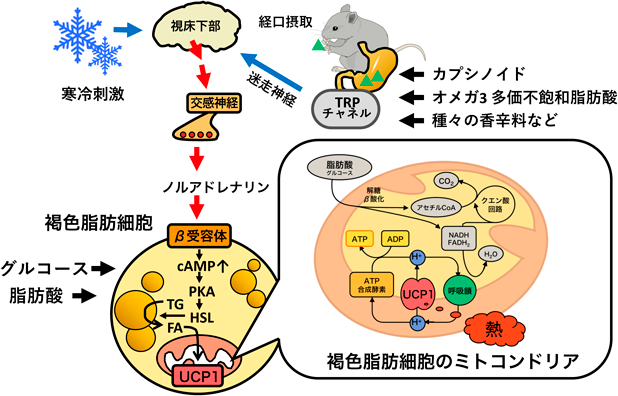
哺乳類には2種類の脂肪組織がある(図1A, B).一般的に内臓脂肪や皮下脂肪と呼ばれている白色脂肪組織(白色脂肪)は,余剰なエネルギーを中性脂肪として蓄え,必要に応じて脂肪酸として全身に供給するエネルギーの貯蔵を担う.加えて,さまざまなアディポカインを合成・分泌して,全身組織の代謝を制御している.褐色脂肪組織(褐色脂肪)は,白色脂肪と同様に中性脂肪を蓄えるが,活性化すると脂肪エネルギーを消費して熱を産生する3, 4).熱産生を担うのは褐色脂肪細胞のミトコンドリア内膜に存在する脱共役タンパク質1(uncoupling protein 1:UCP1)であり,ミトコンドリアにおけるATP合成を脱共役し,そのエネルギーを熱として散逸する(図2).UCP1の活性は交感神経により制御されており,たとえば寒冷刺激は中枢を介して褐色脂肪を支配する交感神経を活性化し,神経終末から放出されるノルアドレナリンが褐色脂肪細胞のβアドレナリン受容体に結合すると,細胞内ではアデニル酸シクラーゼ-cAMP-プロテインキナーゼAといったシグナル伝達経路が活性化され,その下流でホルモン感受性リパーゼが活性化して中性脂肪が分解される.生じた脂肪酸は熱産生の基質として消費されるとともに,直接UCP1を活性化する.この一連の反応により褐色脂肪で熱が産生され(非震え熱産生)寒冷環境においても体温が維持される.
交感神経性刺激が長期にわたれば,UCP1遺伝子発現量や褐色脂肪細胞の数が増加し,全身の熱産生能が増大する.我々は褐色脂肪細胞の数の増加には,前駆細胞の増殖・分化のみならず成熟褐色脂肪細胞の分裂も関わることを報告している5, 6).褐色脂肪は熱産生の基質として脂肪エネルギーを消費するため,褐色脂肪の増加や活性化は体脂肪を減少させる.たとえばマウスにβ3アドレナリン受容体作動薬を長期間投与すると,褐色脂肪の熱産生が増加し,全身のエネルギー消費量が増加して体脂肪が減少する7).
ヒトにおいては寒冷刺激と18F-FDG-PET/CTを組み合わせることにより,活性化した褐色脂肪を検出することができる1)(図1C).PET-CTでの検出は,褐色脂肪細胞−UCP1が活性化すると細胞内へのグルコースの取り込みが増加する代謝特性8)に基づいている.ヒトの褐色脂肪量は体脂肪量と逆相関するので1),ヒトにおいても褐色脂肪が体脂肪量の調節に関わっていると考えられる.
2)ベージュ脂肪細胞
白色脂肪細胞は細胞内に一つの大きな脂肪滴(単房性脂肪滴)を含むのに対し,褐色脂肪細胞は多数の小さな脂肪滴(多房性脂肪滴)を含むという形態の違いがある.一方,慢性的な交感神経性刺激により白色脂肪組織中に多房性脂肪滴を持つ脂肪細胞が新たに誘導される.褐色脂肪組織に存在する褐色脂肪細胞(classical brown adipocyte)と同様の熱産生機能を持つが9),細胞系譜や遺伝子発現パターンに違いがあるため10, 11),brite/beige脂肪細胞として区別されている(本稿ではベージュ脂肪細胞と呼ぶ).
ベージュ脂肪細胞は白色脂肪組織中に存在する前駆細胞が分化して生じると考えられるが,少なくとも一部は白色脂肪細胞からの転換により生じること12, 13)が報告されている.交感神経性刺激がなくなるとマイトファジーによりミトコンドリアが消失して白色脂肪細胞様の形態になる14)ことから,白色脂肪組織中の白色脂肪細胞は形態学的には均一であってもヘテロな集団であり,一部が刺激に応じて形態と機能を変化させているのかもしれない.興味深いことに,成人の褐色脂肪を構成するのはベージュ脂肪細胞,もしくはベージュ脂肪細胞と褐色脂肪細胞(classical brown adipocyte)の混合であることが明らかになっている15, 16).そのため,ベージュ脂肪細胞の誘導メカニズムについて活発に研究が進められており,交感神経性刺激以外にも古くから知られている甲状腺ホルモンに加えて,肝臓から分泌されるfibroblast growth factor 2116)や胆汁酸17),心臓からのナトリウム利尿ペプチド(natriuretic peptide)18)などが誘導因子として報告されている.
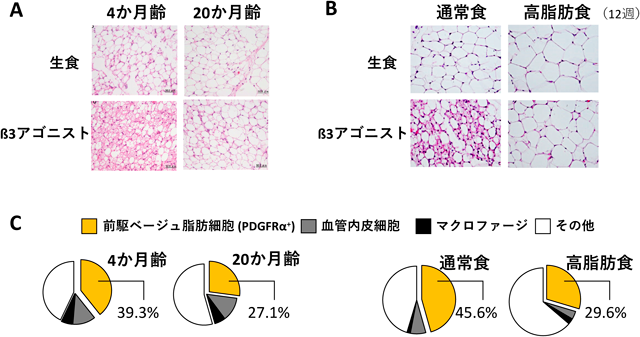
一般に,classical brown adipocyteは新生児期の体温維持に重要であり19),ヒトを含めた多くの動物種において成長とともに減少する.マウスやラットなどの小型動物ではclassical brown adipocyteは生涯維持されるが,ベージュ脂肪細胞の誘導性に加齢性変化があるのかは不明であった.我々はマウスにおいて加齢はß3アドレナリン受容体作動薬CL316,243(CL)によるベージュ脂肪細胞の誘導を著しく低下させることを見いだした(図3A, C)20).加齢マウスの白色脂肪では,ベージュ脂肪細胞への分化能を持つことが報告されている血小板由来増殖因子受容体(platelet-derived growth factor receptor:PDGFR)α陽性細胞21)の数が有意に少なかった.マウスに高脂肪食を与えて食事性肥満を誘導すると,やはりPDGFRα細胞が減少してベージュ脂肪細胞の誘導量が低下した(図3B, C)22).食事性肥満ではPDGFRαのリガンドであるPDGF-Bの遺伝子発現が低下しており,PDGFRシグナルの阻害剤はCL投与によるベージュ脂肪細胞の誘導を減弱させたため,PDGFRαを介した増殖が低下してPDGFRα細胞の数が減少し,ベージュ脂肪細胞の誘導量が低下したと考えられた.
以上のように,ベージュ脂肪細胞の制御には前駆細胞であるPDGFRα細胞の増殖メカニズムの解明が必要であると思われる.PDGFRαのリガンドとなるPDGFにはAからDの4種類があり,ホモ二量体またはヘテロ二量体を形成して機能するが,ベージュ脂肪細胞の誘導には血管内皮細胞が分泌するPDGF-Cのホモ二量体が重要であるという報告や23),PDGFRα細胞とPDGFRβ細胞の数のバランスが重要であること24, 25)などが報告されている.
1)過食による白色脂肪の肥大
脂肪組織は食事により摂取したエネルギーを蓄える貯蔵庫であるため,当然のことながら食事の量や質は脂肪組織に大きな影響を与える.過食や高脂肪の食事による(脂肪)エネルギー摂取量の増大は白色脂肪組織の増大を引き起こし,肥満やメタボリックシンドロームを引き起こす.中性脂肪含量の増大により肥大化した白色脂肪細胞ではアディポカインやリポカインの分泌異常が起こる.たとえば,インスリン感受性を増加させるアディポネクチンの分泌が低下し,インスリン抵抗性を引き起こすTNF-α(tumor necrosis factor-α),血栓の形成を促進するプラスミノゲンアクチベーターインヒビター(plasminogen activator inhibitor 1:PAI-1),血圧上昇を引き起こすアンジオテンシノーゲンなどの分泌が増大する.またケモカインであるmonocyte chemotactic protein 1(MCP1)の発現が増加し,組織中への炎症性M1マクロファージの浸潤を促す.マクロファージからはTNF-αやIL-6などの炎症性サイトカインが分泌されて脂肪組織に低レベルの慢性炎症を引き起こすとともに,インスリン抵抗性を増悪させる.肥満による白色脂肪組織の変化と全身代謝への影響については優れた総説が多くあるのでそれらを参照していただきたい26–29).
2)食事誘発性熱産生と褐色脂肪
食事をとると体温が上昇することを我々は経験的に知っているが,食後には消化・吸収といった活動により熱が産生されることに加えて,自律神経系を介してエネルギー消費量が増大する30).この自律神経系を介した熱産生は食事誘発性熱産生(diet-induced thermogenesis:DIT)と呼ばれ,褐色脂肪が関わると考えられている.1979年にRothwellとStockは,マウスにカフェテリア食という嗜好性の高い食餌を与えると摂取カロリーが増加するがその割には太らないことを報告し,褐色脂肪の熱産生能が増大していることを示した31).長期的なエネルギー摂取の増大に対しエネルギーバランスを保つためのホメオスタシス維持機構として,余剰なエネルギーを熱として放散するいわゆる“luxury consumption”の経路が存在し,褐色脂肪が一定の寄与をすることが示唆された.1981年にGlickらがラットにおいて餌を食べた2時間後には褐色脂肪の熱産生が活性化していることを示し32),1回の食事によっても褐色脂肪の熱産生が誘導されることを示した.この経路の誘導は長期的には体脂肪に影響を及ぼすと考えられるが,実際にUCP1欠損マウスを体温調節のための熱産生を行う必要のない室温30度の温暖環境(thermoneutral condition)で飼育すると,高脂肪食による食事性肥満が誘導されやすいことが報告されている33).ヒトでも,DITは痩せ型の人に比べて肥満者では低いことが報告されており34),肥満の成因の一つであることが想定されている.
ヒトにおいて通常の食事内容でのDITはエネルギー摂取量のおよそ10%であるが35),食事の組成によっても変化し,脂肪では0~3%,炭水化物では5~10%,タンパク質では20~30%がDITとして消費される36).つまり,食事の質はDITに大きな影響を与える.また,DITは昼食や夕食後よりも朝食後に高い37)など食事の時間帯の影響もある.さらに興味深いことに食事の美味しさ(palatability)もDITを規定する要因である.LeBlancとBrondelは1985年に興味深い研究成果を報告している38).8名の女性を2群に分け,一方には“美味しい”食事を,もう一方のグループにはその食事をすべて混合して作製したビスケットとして与えると,美味しい食事を食べた後には熱産生量が20%増加したのに対し,同じカロリーと栄養素を含むにもかかわらずビスケットとして摂取したグループでは12%しか増加しなかった.血中のノエルエピネフリン濃度も美味しい食事を食べたグループでのみ増加しており,美味しさを味わって食べることが交感神経を介してDITを増加させることを示している.同グループはイヌを用いた実験によりさらに詳細な検討を行っている39).イヌに餌を与えると食後40分以内に速やかに起こる熱産生と,その後に穏やかに2時間ほど続く熱産生の二峰性のエネルギー消費量の増大が認められたが,同じ食事内容をチューブを介して直接胃内に入れた場合には前半の熱産生が抑制され,餌を味わって食べるものの胃には入らないように外科的に処置した群では後半の熱産生が抑制された.したがって,前半の熱産生が“美味しさ”を感じることにより生じるDITを,後半の熱産生が消化・吸収に伴う熱産生を示していると考えられる.ラットにおいても,液体食を口からまたはチューブを介して直接胃に与えると,同じ量の餌を摂取したにもかかわらずチューブ食では通常摂取よりも体脂肪が増加し,交感神経活動の指標である褐色脂肪のノルエピネフリン代謝回転が低下する40).
ヒトUCP1遺伝子のプロモーター領域には一塩基多型(−3826 A/G)が存在し41),エネルギー摂取量の減少による体重低下42)や加齢に伴う褐色脂肪の減少43)と関連するが,DITもまたこの遺伝子型に関連する44)ことから,ヒトにおいてもDITに褐色脂肪が一定の寄与をすることが示唆される.一方で,長期の寒冷馴化は寒冷誘導性熱産生を増加させるがDITには影響しないこと45)や,DITがβアドレナリン受容体阻害の影響を受けない46, 47)などの報告もあり,褐色脂肪の寄与について一定の結論が得られていなかった.しかし,近年のFDG-PET/CTを用いたヒト褐色脂肪研究の進展により直接的な検討が可能になった.Hibiらはヒューマンカロリメーターを用いて24時間の代謝変化を測定し,FDG-PET/CTの結果に基づき褐色脂肪の活性が高いグループと低いグループに分けて解析すると,安静時のエネルギー消費量には違いはないものの,食後のエネルギー消費量は褐色脂肪高活性のグループで有意に高いことを見いだした48).したがって,ヒトにおいてもDITに褐色脂肪による熱産生が一定の寄与をしていると考えられる.
1)香辛料
寒冷刺激は温度受容体であるtransient receptor potential(TRP)チャネルを活性化し,感覚神経を介して脳に伝えられて交感神経–褐色脂肪–UCP1の経路を活性化する.TRPチャネルは温度,pH,浸透圧,機械刺激などさまざまな刺激により活性化するが49),TRPファミリーの一つTRPV1は唐辛子の辛み成分であるカプサイシンにより活性化する.カプサイシンを摂取するとエネルギー消費量が増加し,慢性的に摂取すると体脂肪が減少することがヒトやマウスの実験で示されている50–53).褐色脂肪への作用については報告が少ないものの,UCP1を増加させる可能性がラットを用いた実験により示されている54).
カプサイシンは強い辛みを示すため肥満対策に利用するには適していないが,カプサイシンの類似体であるカプシノイドは辛みが少なく(カプサイシンの1000分の1),カプサイシンと同様にTRPV1チャネルを活性化する55).カプシノイドはカプシエイト,ジヒドロカプシエイト,ノルジヒドロカプシエイトからなり,マウスやヒトにおいて,摂取するとエネルギー消費量が亢進し56, 57),長期摂取により肥満が軽減されることが示されている58, 59).カプシノイドの体脂肪減少作用はTRPV1欠損マウスでは認められず,胃からの迷走神経切除によっても消失することから56),摂取したカプシノイドは胃のTRPV1チャネルを活性化し,迷走神経を介して脳に情報が伝えられると考えられる.我々は,マウスにおいてカプシノイドは高脂肪食による食事性肥満を抑制することや,UCP1欠損マウスでは効果が認められないことを明らかにしている60).ヒトにおいてもカプシノイド摂取による褐色脂肪の増量と寒冷誘導性熱産生の増加が認められている61).これらの事実から,カプシノイドの体脂肪減少作用には褐色脂肪が関わると結論できる.
このように,カプシノイドは胃のTRPV1チャネル–迷走神経–脳–交感神経–褐色脂肪という一連の経路を活性化してエネルギー消費量を亢進させ,肥満を軽減する(図2).カプシノイド以外にも,緑茶の渋み成分であるカテキンや生姜の辛味成分であるジンゲロール,ジンゲロン,ショウガオール,胡椒の辛味成分であるピペリンなどもTRPV1チャネルを介して同様の経路を活性化する可能性が示されている62).加えて,TRPA1チャネルに対してアゴニスト作用を示すマスタードやわさびに含まれるアリルイソチオシアネートやシナモンに含まれるシンナムアルデヒドが褐色脂肪を活性化またはベージュ脂肪細胞を誘導して体脂肪を減少させることがマウスを用いた実験により示されており63–65),これら香辛料による褐色脂肪の活性化は有効な肥満対策につながることが期待されている.
2)オメガ3多価不飽和脂肪酸(omega-3 polyunsaturated fatty acids:ω3 PUFAs)
魚油に含まれるエイコサペンタエン酸(eicosapentaenoic acid:EPA)やドコサヘキサエン酸(docosahexaenoic acid:DHA)といったω3 PUFAsは,抗炎症作用66)や心血管保護作用67)などに加えて,食事性肥満を抑制することが知られている.そのメカニズムとして白色脂肪での炎症抑制68)や脂肪酸酸化の促進69)が提唱されている他,褐色脂肪の関与を示す報告も多い.たとえば,マウスやラットにEPAやDHAを与えると,褐色脂肪UCP1発現量の増加70)や,UCP1活性の指標であるGDP結合能の増加と褐色脂肪の過形成71),エネルギー消費の亢進による食事性肥満の抑制72)がみられる.一方で,魚油やEPAによる食事性肥満の抑制はUCP1欠損マウスでも認められ73, 74),Ca2+サイクリング(小胞体や筋小胞体からのCa2+遊離と再取り込みを繰り返すサイクル.エネルギーを浪費する無益回路の一つ)を介した代償的な熱産生経路75)が誘導される73)ことが示されており,褐色脂肪の寄与については不明な点が多い.
ω3 PUFAsの体脂肪減少作用を示す分子メカニズムとしてはいくつかの報告がある.KimらはマウスにDHAやEPAを与えると全身のエネルギー消費量が亢進し,同時に褐色脂肪と白色脂肪でのUCP1発現量や交感神経活動の増加が認められたが,TRPV1欠損マウスではこれらの作用が消失することを示した76).一方,ω3 PUFAsによる褐色/ベージュ脂肪細胞でのUCP1発現誘導が脂肪酸受容体であるGPR120/FFAR4を介すること77),GPR120欠損マウスでは消失すること78)が示されている.さらに,ω3 PUFAsの代謝産物であるoxylipinがGタンパク質共役受容体や核内受容体であるペルオキシソーム増殖剤活性化受容体(peroxisome proliferator-activated receptor:PPAR)を介して作用を示す可能性もあり79),EPA由来の12-HEPEが寒冷刺激により褐色脂肪より分泌され,白色脂肪や筋肉のグルコース利用を増加させることが報告されている80).
ヒトにおいては,ω3 PUFAsの摂取はDITを増加させる81, 82)ことが示されているが,その作用に褐色脂肪が関与しているかについてはまだ不明である.
3)ビタミンAとビタミンD
畜産分野では肉牛の脂肪交雑(筋間脂肪)を増やすために肥育期間中に餌中のビタミンAの制限が行われているが,ビタミンAは脂肪細胞の分化を抑制する.ビタミンA(レチノール,レチナール,レチノイン酸とその類縁化合物)は目の発達,皮膚や粘膜の恒常性維持,細胞の分化と個体発生などに関わる多彩な生理作用を示す脂溶性ビタミンである.ヒトを含め動物は動物性食品からレチノールや脂肪酸と結合した脂肪酸エステル(レニチルエステル)として,または植物性食品からビタミンA前駆体であるβカロテンなどのカロテノイドとして摂取している.カロテノイドは小腸粘膜で酵素的にレチノールに転換される.レチノールはエステル化されてキロミクロンに取り込まれ,リンパ管と門脈を経て肝臓に運ばれ,再びレチノールに分解されレチノール結合タンパク質(retinol-binding protein:RBP)に結合して血中を運搬される.細胞内に入るとレチナールへ,続いてレチノイン酸(retinoic acid:RA)へと変換される.細胞内ではRAは細胞性レチノイン酸結合タンパク質(cellular retinoic acid-binding protein:CRABP)により核内に運ばれ,all-trans-RAはレチノイン酸受容体(retinoic acid receptor:RAR)と,9-cis-RAはRARまたはレチノイドX受容体(retinoid X receptor:RXR)と結合し,RARとRXRはヘテロ二量体を形成して遺伝子プロモーターのレチノイン酸応答エレメント(retinoic acid response element:RARE)に結合して発現を制御する83).all-trans-RAは脂肪酸結合タンパク質(fatty acid-binding protein:FABP)5により核内に運ばれてPPARδのリガンドとしても機能する84).
RAは脂肪細胞の分化を抑制するが,その作用は分化初期に限定されている85).分子メカニズムとして,前駆脂肪細胞ではRAはCRABPに運ばれてRARのリガンドとなり前駆細胞特異的な遺伝子発現を促進することで分化を抑制することが示されている86).脂肪細胞の分化の初期に発現が増加するCCA AT/enhancer binding protein α(C/EBPα)がCRABP発現を抑制するとともに,FABP5とPPARδの発現が増加して分化を進める87).in vivoでは,マウスにおいてビタミンAの欠乏は白色脂肪組織を増大させ,反対にall-trans-RAの投与は体脂肪を減少させる88).前述のとおり,ウシでは低ビタミンA食は筋間脂肪を増加させる89).
ラットとマウスのUCP1遺伝子プロモーターにはRAREがあり90),実際RA処置や高ビタミンA食は褐色脂肪細胞のUCP1発現を増加させることがin vitroおよびin vivoにおいて示されている91, 92).ヒトUCP1遺伝子もRAに応答してプロモーター活性が上昇する93).興味深いことに,RAは血管内皮細胞に作用して血管内皮増殖因子の発現を増加させ,前駆ベージュ脂肪細胞であるPDGFRα細胞数を増加させることが報告されている94).
ビタミンAと同じく脂溶性ビタミンであるビタミンDは核内に存在するビタミンD受容体(vitamin D receptor:VDR)を介して脂肪分化を抑制する.その抑制作用はやはり分化初期にのみ生じ,初期分化に必須の転写因子であるC/EBPαやPPARγ発現の抑制やPPARγ活性の抑制による95).ヒトにおいてビタミンDの欠乏は肥満に関連すること96),ビタミンD結合タンパク質97)やビタミンD受容体98)の一塩基多型は肥満や2型糖尿病と関連することが報告されている.一方で,VDR欠損マウスは体脂肪が少なく食事性肥満にも抵抗性であること99, 100),脂肪細胞特異的VDR過剰発現マウスでは,摂食量は変わらないにもかかわらず肥満する101)など,VDRによる白色脂肪分化への影響は必ずしも明確ではない.なお,ヒトのUCP1遺伝子のプロモーター領域にはビタミンD応答配列(vitamin D response element:VDRE)があり,VDRにより転写が抑制される102)が,この抑制作用はリガンド非依存的であることが示されており,ビタミンDが白色/褐色脂肪に与える影響についてはさらなる検討が必要である.
ここまで食が脂肪組織に与える影響についてまとめてきたが,脂肪組織は食欲の調節に重要な組織である.遺伝性肥満のob/obマウスの原因遺伝子として単離・同定されたアディポカインであるレプチンは,摂食中枢である視床下部に作用して食欲を抑制するとともに交感神経活動を亢進させてエネルギー消費を増強させる.肥満すると血中レプチン濃度が慢性的に増加してレプチンの作用障害(レプチン抵抗性)が誘導され,多量のレプチンが存在するにもかかわらず作用を示さないレプチン抵抗性となる.レプチン抵抗性の発症機序としてレプチンの脳内への運搬が低下することや103),SOCS3(suppressor of cytokine signaling 3)をはじめとした負の制御因子によるレプチンシグナル伝達の減弱などが原因104, 105)とされている.
食欲の制御について古くからいくつかの仮説が提唱され,後に分子機序が明らかにされてきた.たとえば,体脂肪量を一定に保つように食欲が調節されているとする脂肪定常説はレプチンの発見により,血糖値を保つように調節されているとする糖定常説は,脳内のグルコース感受性神経が血糖値レベルを感知して満腹感を引き起こす経路により証明された.これらの仮説に加えて提唱されてきたのが温度定常説である106).体温があるセットポイントより高くなると食欲が低下し,セットポイントより低くなると食欲が増加するという仮説である.体温が上がると食欲が低下することを経験する人は少なくないと思うが,そのような体温による食欲調節があるのかは不明である.
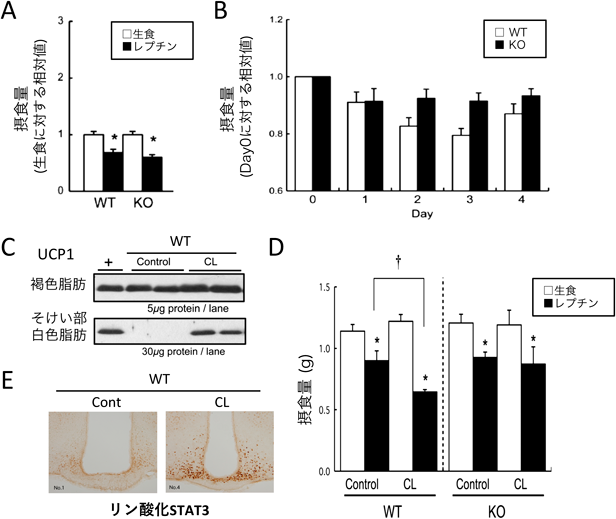
我々は,UCP1がレプチンのエネルギー消費亢進作用のみならず107),摂食抑制作用にも関わる可能性を見いだした108).マウスにレプチンを投与すると摂食量が減少するが,慢性的なレプチン刺激を与えると徐々に摂食抑制作用が増強する(図4).UCP1欠損マウスではそのような増強は認められないため,何らかのUCP1依存的な仕組みによりレプチン感受性が増大すると考えられた.慢性的なレプチン刺激は白色脂肪組織のUCP1発現量を増加させたため,マウスにβ3アゴニスト(CL)を3日間投与して白色脂肪組織中にベージュ脂肪細胞を誘導してからレプチンを投与すると,レプチン感受性が増加することがわかった.UCP1欠損マウスではCL投与によるレプチン感受性の変化は認められなかった.これらの結果は,ベージュ脂肪細胞の誘導により全身の熱産生能が亢進すると,レプチン感受性が増加して摂食量が減少することを示している.Yamadaらはアデノウイルスベクターにより白色脂肪組織にUCP1を発現させると,その情報が求心性神経を介して中枢に伝えられてインスリンやレプチン感受性を亢進させることを報告しており109),アディポカインなど液性シグナルに加えて求心性神経シグナルもまた脂肪組織からの情報発信に寄与していると考えられる.これらの結果は,体温や熱産生による食欲調節の可能性を示している.
引用文献References
1) Saito, M., Okamatsu-Ogura, Y., Matsushita, M., Watanabe, K., Yoneshiro, T., Nio-Kobayashi, J., Iwanaga, T., Miyagawa, M., Kameya, T., Nakada, K., et al. (2009) High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes, 58, 1526–1531.
2) Nedergaard, J., Bengtsson, T., & Cannon, B. (2007) Unexpected evidence for active brown adipose tissue in adult humans. Am. J. Physiol. Endocrinol. Metab., 293, E444–E452.
3) Cannon, B. & Nedergaard, J. (2004) Brown adipose tissue: function and physiological significance. Physiol. Rev., 84, 277–359.
4) 斉藤昌之,大野秀樹編(2013)ここまでわかった燃える褐色脂肪の不思議,有限会社ナップ.
5) Fukano, K., Okamatsu-Ogura, Y., Tsubota, A., Nio-Kobayashi, J., & Kimura, K. (2016) Cold Exposure Induces Proliferation of Mature Brown Adipocyte in a β3-Adrenergic Receptor-Mediated Pathway. PLoS One, 11, e0166579.
6) Okamatsu-Ogura, Y., Fukano, K., Tsubota, A., Nio-Kobayashi, J., Nakamura, K., Morimatsu, M., Sakaue, H., Saito, M., & Kimura, K. (2017) Cell-cycle arrest in mature adipocytes impairs BAT development but not WAT browning, and reduces adaptive thermogenesis in mice. Sci. Rep., 7, 6648.
7) Inokuma, K., Okamatsu-Ogura, Y., Omachi, A., Matsushita, Y., Kimura, K., Yamashita, H., & Saito, M. (2006) Indispensable role of mitochondrial UCP1 for antiobesity effect of beta3-adrenergic stimulation. Am. J. Physiol. Endocrinol. Metab., 290, E1014–E1021.
8) Inokuma, K., Ogura-Okamatsu, Y., Toda, C., Kimura, K., Yamashita, H., & Saito, M. (2005) Uncoupling protein 1 is necessary for norepinephrine-induced glucose utilization in brown adipose tissue. Diabetes, 54, 1385–1391.
9) Okamatsu-Ogura, Y., Fukano, K., Tsubota, A., Uozumi, A., Terao, A., Kimura, K., & Saito, M. (2013) Thermogenic ability of uncoupling protein 1 in beige adipocytes in mice. PLoS One, 8, e84229.
10) Wu, J., Bostrom, P., Sparks, L.M., Ye, L., Choi, J.H., Giang, A.H., Khandekar, M., Virtanen, K.A., Nuutila, P., Schaart, G., et al. (2012) Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell, 150, 366–376.
11) Seale, P., Bjork, B., Yang, W., Kajimura, S., Chin, S., Kuang, S., Scime, A., Devarakonda, S., Conroe, H.M., Erdjument-Bromage, H., et al. (2008) PRDM16 controls a brown fat/skeletal muscle switch. Nature, 454, 961–967.
12) Frontini, A. & Cinti, S. (2010) Distribution and development of brown adipocytes in the murine and human adipose organ. Cell Metab., 11, 253–256.
13) Rosenwald, M., Perdikari, A., Rulicke, T., & Wolfrum, C. (2013) Bi-directional interconversion of brite and white adipocytes. Nat. Cell Biol., 15, 659–667.
14) Altshuler-Keylin, S., Shinoda, K., Hasegawa, Y., Ikeda, K., Hong, H., Kang, Q., Yang, Y., Perera, R.M., Debnath, J., & Kajimura, S. (2016) Beige Adipocyte Maintenance Is Regulated by Autophagy-Induced Mitochondrial Clearance. Cell Metab., 24, 402–419.
15) Lidell, M.E., Betz, M.J., Dahlqvist Leinhard, O., Heglind, M., Elander, L., Slawik, M., Mussack, T., Nilsson, D., Romu, T., Nuutila, P., et al. (2013) Evidence for two types of brown adipose tissue in humans. Nat. Med., 19, 631–634.
16) Cypess, A.M., White, A.P., Vernochet, C., Schulz, T.J., Xue, R., Sass, C.A., Huang, T.L., Roberts-Toler, C., Weiner, L.S., Sze, C., et al. (2013) Anatomical localization, gene expression profiling and functional characterization of adult human neck brown fat. Nat. Med., 19, 635–639.
17) Worthmann, A., John, C., Ruhlemann, M.C., Baguhl, M., Heinsen, F.A., Schaltenberg, N., Heine, M., Schlein, C., Evangelakos, I., Mineo, C., et al. (2017) Cold-induced conversion of cholesterol to bile acids in mice shapes the gut microbiome and promotes adaptive thermogenesis. Nat. Med., 23, 839–849.
18) Bordicchia, M., Liu, D., Amri, E.Z., Ailhaud, G., Dessi-Fulgheri, P., Zhang, C., Takahashi, N., Sarzani, R., & Collins, S. (2012) Cardiac natriuretic peptides act via p38 MAPK to induce the brown fat thermogenic program in mouse and human adipocytes. J. Clin. Invest., 122, 1022–1036.
19) Tsubota, A., Okamatsu-Ogura, Y., Bariuan, J.V., Mae, J., Matsuoka, S., Nio-Kobayashi, J., & Kimura, K. (2019) Role of brown adipose tissue in body temperature control during the early postnatal period in Syrian hamsters and mice. J. Vet. Med. Sci., 81, 1461–1467.
20) Shin, W., Okamatsu-Ogura, Y., Machida, K., Tsubota, A., Nio-Kobayashi, J., & Kimura, K. (2017) Impaired adrenergic agonist-dependent beige adipocyte induction in aged mice. Obesity (Silver Spring), 25, 417–423.
21) Lee, Y.H., Petkova, A.P., Mottillo, E.P., & Granneman, J.G. (2012) In vivo identification of bipotential adipocyte progenitors recruited by β3-adrenoceptor activation and high-fat feeding. Cell Metab., 15, 480–491.
22) Shin, W., Okamatsu-Ogura, Y., Matsuoka, S., Tsubota, A., & Kimura, K. (2019) Impaired adrenergic agonist-dependent beige adipocyte induction in obese mice. J. Vet. Med. Sci., 81, 799–807.
23) Seki, T., Hosaka, K., Lim, S., Fischer, C., Honek, J., Yang, Y., Andersson, P., Nakamura, M., Naslund, E., Yla-Herttuala, S., et al. (2016) Endothelial PDGF-CC regulates angiogenesis-dependent thermogenesis in beige fat. Nat. Commun., 7, 12152.
24) Gao, Z., Daquinag, A.C., Su, F., Snyder, B., & Kolonin, M.G. (2018). Development, 145.
25) Sun, C., Sakashita, H., Kim, J., Tang, Z., Upchurch, G.M., Yao, L., Berry, W.L., Griffin, T.M., & Olson, L.E. (2020) Mosaic Mutant Analysis Identifies PDGFRα/PDGFRβ as Negative Regulators of Adipogenesis. Cell Stem Cell, 26, 707–721.
26) Sun, K., Kusminski, C.M., & Scherer, P.E. (2011) Adipose tissue remodeling and obesity. J. Clin. Invest., 121, 2094–2101.
27) Kusminski, C.M., Bickel, P.E., & Scherer, P.E. (2016) Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat. Rev. Drug Discov., 15, 639–660.
28) Kita, S., Maeda, N., & Shimomura, I. (2019) Interorgan communication by exosomes, adipose tissue, and adiponectin in metabolic syndrome. J. Clin. Invest., 129, 4041–4049.
29) Kahn, C.R., Wang, G., & Lee, K.Y. (2019) Altered adipose tissue and adipocyte function in the pathogenesis of metabolic syndrome. J. Clin. Invest., 129, 3990–4000.
30) Tappy, L. (1996) Thermic effect of food and sympathetic nervous system activity in humans. Reprod. Nutr. Dev., 36, 391–397.
31) Rothwell, N.J. & Stock, M.J. (1979) A role for brown adipose tissue in diet-induced thermogenesis. Nature, 281, 31–35.
32) Glick, Z., Teague, R.J., & Bray, G.A. (1981) Brown adipose tissue: thermic response increased by a single low protein, high carbohydrate meal. Science, 213, 1125–1127.
33) Feldmann, H.M., Golozoubova, V., Cannon, B., & Nedergaard, J. (2009) UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metab., 9, 203–209.
34) de Jonge, L. & Bray, G.A. (1997) The thermic effect of food and obesity: a critical review. Obes. Res., 5, 622–631.
35) Westerterp, K.R. (2004) Diet induced thermogenesis. Nutr. Metab. (Lond.), 1, 5.
36) Acheson, K.J. (1993) Influence of autonomic nervous system on nutrient-induced thermogenesis in humans. Nutrition, 9, 373–380.
37) Romon, M., Edme, J.L., Boulenguez, C., Lescroart, J.L., & Frimat, P. (1993) Circadian variation of diet-induced thermogenesis. Am. J. Clin. Nutr., 57, 476–480.
38) LeBlanc, J. & Brondel, L. (1985) Role of palatability on meal-induced thermogenesis in human subjects. Am. J. Physiol., 248, E333–E336.
39) Diamond, P., Brondel, L., & LeBlanc, J. (1985) Palatability and postprandial thermogenesis in dogs. Am. J. Physiol., 248, E75–E79.
40) Saito, M., Minokoshi, Y., & Shimazu, T. (1989) Metabolic and sympathetic nerve activities of brown adipose tissue in tube-fed rats. Am. J. Physiol., 257, E374–E378.
41) Esterbauer, H., Oberkofler, H., Liu, Y.M., Breban, D., Hell, E., Krempler, F., & Patsch, W. (1998) Uncoupling protein-1 mRNA expression in obese human subjects: the role of sequence variations at the uncoupling protein-1 gene locus. J. Lipid Res., 39, 834–844.
42) Nagai, N., Sakane, N., Kotani, K., Hamada, T., Tsuzaki, K., & Moritani, T. (2011) Uncoupling protein 1 gene -3826 A/G polymorphism is associated with weight loss on a short-term, controlled-energy diet in young women. Nutr. Res., 31, 255–261.
43) Yoneshiro, T., Ogawa, T., Okamoto, N., Matsushita, M., Aita, S., Kameya, T., Kawai, Y., Iwanaga, T., & Saito, M. (2013) Impact of UCP1 and β3AR gene polymorphisms on age-related changes in brown adipose tissue and adiposity in humans. Int. J. Obes., 37, 993–998.
44) Nagai, N., Sakane, N., Ueno, L.M., Hamada, T., & Moritani, T. (2003) The -3826 A→G variant of the uncoupling protein-1 gene diminishes postprandial thermogenesis after a high fat meal in healthy boys. J. Clin. Endocrinol. Metab., 88, 5661–5667.
45) Peterson, C.M., Lecoultre, V., Frost, E.A., Simmons, J., Redman, L.M., & Ravussin, E. (2016) The thermogenic responses to overfeeding and cold are differentially regulated. Obesity (Silver Spring), 24, 96–101.
46) Thorne, A. & Wahren, J. (1989) Beta-adrenergic blockade does not influence the thermogenic response to a mixed meal in man. Clin. Physiol., 9, 321–332.
47) Welle, S. & Campbell, R.G. (1983) Stimulation of thermogenesis by carbohydrate overfeeding. Evidence against sympathetic nervous system mediation. J. Clin. Invest., 71, 916–925.
48) Hibi, M., Oishi, S., Matsushita, M., Yoneshiro, T., Yamaguchi, T., Usui, C., Yasunaga, K., Katsuragi, Y., Kubota, K., Tanaka, S., et al. (2016) Brown adipose tissue is involved in diet-induced thermogenesis and whole-body fat utilization in healthy humans. Int. J. Obes., 40, 1655–1661.
49) Uchida, K., Dezaki, K., Yoneshiro, T., Watanabe, T., Yamazaki, J., Saito, M., Yada, T., Tominaga, M., & Iwasaki, Y. (2017) Involvement of thermosensitive TRP channels in energy metabolism. J. Physiol. Sci., 67, 549–560.
50) Kawada, T., Watanabe, T., Takaishi, T., Tanaka, T., & Iwai, K. (1986) Capsaicin-induced beta-adrenergic action on energy metabolism in rats: influence of capsaicin on oxygen consumption, the respiratory quotient, and substrate utilization. Proc. Soc. Exp. Biol. Med., 183, 250–256.
51) Watanabe, T., Kawada, T., Kurosawa, M., Sato, A., & Iwai, K. (1988) Adrenal sympathetic efferent nerve and catecholamine secretion excitation caused by capsaicin in rats. Am. J. Physiol., 255, E23–E27.
52) Yoshioka, M., St-Pierre, S., Suzuki, M., & Tremblay, A. (1998) Effects of red pepper added to high-fat and high-carbohydrate meals on energy metabolism and substrate utilization in Japanese women. Br. J. Nutr., 80, 503–510.
53) Kawada, T., Hagihara, K., & Iwai, K. (1986) Effects of capsaicin on lipid metabolism in rats fed a high fat diet. J. Nutr., 116, 1272–1278.
54) Kawada, T., Sakabe, S., Aoki, N., Watanabe, T., Higeta, K., Iwai, K., & Sugimoto, E. (1991) Intake of sweeteners and pungent ingredients increases the thermogenin content in brown adipose tissue of rat. J. Agric. Food Chem., 39, 651–654.
55) Kobata, K., Sutoh, K., Todo, T., Yazawa, S., Iwai, K., & Watanabe, T. (1999) Nordihydrocapsiate, a new capsinoid from the fruits of a nonpungent pepper, capsicum annuum. J. Nat. Prod., 62, 335–336.
56) Kawabata, F., Inoue, N., Masamoto, Y., Matsumura, S., Kimura, W., Kadowaki, M., Higashi, T., Tominaga, M., Inoue, K., & Fushiki, T. (2009) Non-pungent capsaicin analogs (capsinoids) increase metabolic rate and enhance thermogenesis via gastrointestinal TRPV1 in mice. Biosci. Biotechnol. Biochem., 73, 2690–2697.
57) Inoue, N., Matsunaga, Y., Satoh, H., & Takahashi, M. (2007) Enhanced energy expenditure and fat oxidation in humans with high BMI scores by the ingestion of novel and non-pungent capsaicin analogues (capsinoids). Biosci. Biotechnol. Biochem., 71, 380–389.
58) Ohyama, K., Nogusa, Y., Suzuki, K., Shinoda, K., Kajimura, S., & Bannai, M. (2015) A combination of exercise and capsinoid supplementation additively suppresses diet-induced obesity by increasing energy expenditure in mice. Am. J. Physiol. Endocrinol. Metab., 308, E315–E323.
59) Snitker, S., Fujishima, Y., Shen, H., Ott, S., Pi-Sunyer, X., Furuhata, Y., Sato, H., & Takahashi, M. (2009) Effects of novel capsinoid treatment on fatness and energy metabolism in humans: possible pharmacogenetic implications. Am. J. Clin. Nutr., 89, 45–50.
60) Okamatsu-Ogura, Y., Tsubota, A., Ohyama, K., Nogusa, Y., Saito, M., & Kimura, K. (2015) Capsinoids suppress diet-induced obesity through uncoupling protein 1-dependent mechanism in mice. J. Funct. Foods, 19, 1–9.
61) Yoneshiro, T., Aita, S., Matsushita, M., Kayahara, T., Kameya, T., Kawai, Y., Iwanaga, T., & Saito, M. (2013) Recruited brown adipose tissue as an antiobesity agent in humans. J. Clin. Invest., 123, 3404–3408.
62) Saito, M., Matsushita, M., Yoneshiro, T., & Okamatsu-Ogura, Y. (2020) Brown Adipose Tissue, Diet-Induced Thermogenesis, and Thermogenic Food Ingredients: From Mice to Men. Front. Endocrinol. (Lausanne), 11, 222.
63) Tamura, Y., Iwasaki, Y., Narukawa, M., & Watanabe, T. (2012) Ingestion of cinnamaldehyde, a TRPA1 agonist, reduces visceral fats in mice fed a high-fat and high-sucrose diet. J. Nutr. Sci. Vitaminol. (Tokyo), 58, 9–13.
64) Zuo, J., Zhao, D., Yu, N., Fang, X., Mu, Q., Ma, Y., Mo, F., Wu, R., Ma, R., Wang, L., et al. (2017) Cinnamaldehyde Ameliorates Diet-Induced Obesity in Mice by Inducing Browning of White Adipose Tissue. Cell. Physiol. Biochem., 42, 1514–1525.
65) Masamoto, Y., Kawabata, F., & Fushiki, T. (2009) Intragastric administration of TRPV1, TRPV3, TRPM8, and TRPA1 agonists modulates autonomic thermoregulation in different manners in mice. Biosci. Biotechnol. Biochem., 73, 1021–1027.
66) Calder, P.C. (2013) Omega-3 polyunsaturated fatty acids and inflammatory processes: nutrition or pharmacology? Br. J. Clin. Pharmacol., 75, 645–662.
67) Sakamoto, A., Saotome, M., Iguchi, K., & Maekawa, Y. (2019). Int. J. Mol. Sci., 20, .
68) Kalupahana, N.S., Claycombe, K., Newman, S.J., Stewart, T., Siriwardhana, N., Matthan, N., Lichtenstein, A.H., & Moustaid-Moussa, N. (2010) Eicosapentaenoic acid prevents and reverses insulin resistance in high-fat diet-induced obese mice via modulation of adipose tissue inflammation. J. Nutr., 140, 1915–1922.
69) LeMieux, M.J., Kalupahana, N.S., Scoggin, S., & Moustaid-Moussa, N. (2015) Eicosapentaenoic acid reduces adipocyte hypertrophy and inflammation in diet-induced obese mice in an adiposity-independent manner. J. Nutr., 145, 411–417.
70) Takahashi, Y. & Ide, T. (2000) Dietary n-3 fatty acids affect mRNA level of brown adipose tissue uncoupling protein 1, and white adipose tissue leptin and glucose transporter 4 in the rat. Br. J. Nutr., 84, 175–184.
71) Oudart, H., Groscolas, R., Calgari, C., Nibbelink, M., Leray, C., Le Maho, Y., & Malan, A. (1997) Brown fat thermogenesis in rats fed high-fat diets enriched with n-3 polyunsaturated fatty acids. Int. J. Obes. Relat. Metab. Disord., 21, 955–962.
72) Pahlavani, M., Razafimanjato, F., Ramalingam, L., Kalupahana, N.S., Moussa, H., Scoggin, S., & Moustaid-Moussa, N. (2017) Eicosapentaenoic acid regulates brown adipose tissue metabolism in high-fat-fed mice and in clonal brown adipocytes. J. Nutr. Biochem., 39, 101–109.
73) Oliveira, T.E., Castro, E., Belchior, T., Andrade, M.L., Chaves-Filho, A.B., Peixoto, A.S., Moreno, M.F., Ortiz-Silva, M., Moreira, R.J., Inague, A., et al. (2019) Fish Oil Protects Wild Type and Uncoupling Protein 1-Deficient Mice from Obesity and Glucose Intolerance by Increasing Energy Expenditure. Mol. Nutr. Food Res., 63, e1800813.
74) Pahlavani, M., Ramalingam, L., Miller, E.K., Scoggin, S., Menikdiwela, K.R., Kalupahana, N.S., Festuccia, W.T., & Moustaid-Moussa, N. (2019) Eicosapentaenoic Acid Reduces Adiposity, Glucose Intolerance and Increases Oxygen Consumption Independently of Uncoupling Protein 1. Mol. Nutr. Food Res., 63, e1800821.
75) Ikeda, K., Kang, Q., Yoneshiro, T., Camporez, J.P., Maki, H., Homma, M., Shinoda, K., Chen, Y., Lu, X., Maretich, P., et al. (2017) UCP1-independent signaling involving SERCA2b-mediated calcium cycling regulates beige fat thermogenesis and systemic glucose homeostasis. Nat. Med., 23, 1454–1465.
76) Kim, M., Goto, T., Yu, R., Uchida, K., Tominaga, M., Kano, Y., Takahashi, N., & Kawada, T. (2015) Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci. Rep., 5, 18013.
77) Kim, J., Okla, M., Erickson, A., Carr, T., Natarajan, S.K., & Chung, S. (2016) Eicosapentaenoic Acid Potentiates Brown Thermogenesis through FFAR4-dependent Up-regulation of miR-30b and miR-378. J. Biol. Chem., 291, 20551–20562.
78) Quesada-Lopez, T., Cereijo, R., Turatsinze, J.V., Planavila, A., Cairo, M., Gavalda-Navarro, A., Peyrou, M., Moure, R., Iglesias, R., Giralt, M., et al. (2016) The lipid sensor GPR120 promotes brown fat activation and FGF21 release from adipocytes. Nat. Commun., 7, 13479.
79) Barquissau, V., Ghandour, R.A., Ailhaud, G., Klingenspor, M., Langin, D., Amri, E.Z., & Pisani, D.F. (2017) Control of adipogenesis by oxylipins, GPCRs and PPARs. Biochimie, 136, 3–11.
80) Leiria, L.O., Wang, C.H., Lynes, M.D., Yang, K., Shamsi, F., Sato, M., Sugimoto, S., Chen, E.Y., Bussberg, V., Narain, N.R., et al. (2019) 12-Lipoxygenase Regulates Cold Adaptation and Glucose Metabolism by Producing the Omega-3 Lipid 12-HEPE from Brown Fat. Cell Metab., 30, 768–783.
81) Clevenger, H.C., Kozimor, A.L., Paton, C.M., & Cooper, J.A. (2014) Acute effect of dietary fatty acid composition on postprandial metabolism in women. Exp. Physiol., 99, 1182–1190.
82) Cisneros, L.C.V., Moreno, A.G.M., Lopez-Espinoza, A., & Espinoza-Gallardo, A.C. (2019) Effect of the fatty acid composition of meals on postprandial energy expenditure: a systematic review. Rev. Assoc. Med. Bras., 65, 1022–1031.
83) Villarroya, F., Giralt, M., & Iglesias, R. (1999) Retinoids and adipose tissues: metabolism, cell differentiation and gene expression. Int. J. Obes. Relat. Metab. Disord., 23, 1–6.
84) Berry, D.C. & Noy, N. (2009) All-trans-retinoic acid represses obesity and insulin resistance by activating both peroxisome proliferation-activated receptor beta/delta and retinoic acid receptor. Mol. Cell. Biol., 29, 3286–3296.
85) Kuri-Harcuch, W. (1982) Differentiation of 3T3-F442A cells into adipocytes is inhibited by retinoic acid. Differentiation, 23, 164–169.
86) Berry, D.C., DeSantis, D., Soltanian, H., Croniger, C.M., & Noy, N. (2012) Retinoic acid upregulates preadipocyte genes to block adipogenesis and suppress diet-induced obesity. Diabetes, 61, 1112–1121.
87) Berry, D.C., Soltanian, H., & Noy, N. (2010) Repression of cellular retinoic acid-binding protein II during adipocyte differentiation. J. Biol. Chem., 285, 15324–15332.
88) Ribot, J., Felipe, F., Bonet, M.L., & Palou, A. (2001) Changes of adiposity in response to vitamin A status correlate with changes of PPAR gamma 2 expression. Obes. Res., 9, 500–509.
89) Gorocica-Buenfil, M.A., Fluharty, F.L., Bohn, T., Schwartz, S.J., & Loerch, S.C. (2007) Effect of low vitamin A diets with high-moisture or dry corn on marbling and adipose tissue fatty acid composition of beef steers. J. Anim. Sci., 85, 3355–3366.
90) Alvarez, R., de Andres, J., Yubero, P., Vinas, O., Mampel, T., Iglesias, R., Giralt, M., & Villarroya, F. (1995) A novel regulatory pathway of brown fat thermogenesis. Retinoic acid is a transcriptional activator of the mitochondrial uncoupling protein gene. J. Biol. Chem., 270, 5666–5673.
91) Puigserver, P., Vazquez, F., Bonet, M.L., Pico, C., & Palou, A. (1996) In vitro and in vivo induction of brown adipocyte uncoupling protein (thermogenin) by retinoic acid. Biochem. J., 317, 827–833.
92) Felipe, F., Bonet, M.L., Ribot, J., & Palou, A. (2003) Up-regulation of muscle uncoupling protein 3 gene expression in mice following high fat diet, dietary vitamin A supplementation and acute retinoic acid-treatment. Int. J. Obes. Relat. Metab. Disord., 27, 60–69.
93) del Mar Gonzalez-Barroso, M., Pecqueur, C., Gelly, C., Sanchis, D., Alves-Guerra, M.C., Bouillaud, F., Ricquier, D., & Cassard-Doulcier, A.M. (2000) Transcriptional activation of the human ucp1 gene in a rodent cell line. Synergism of retinoids, isoproterenol, and thiazolidinedione is mediated by a multipartite response element. J. Biol. Chem., 275, 31722–31732.
94) Wang, B., Fu, X., Liang, X., Deavila, J.M., Wang, Z., Zhao, L., Tian, Q., Zhao, J., Gomez, N.A., Trombetta, S.C., et al. (2017) Retinoic acid induces white adipose tissue browning by increasing adipose vascularity and inducing beige adipogenesis of PDGFRα+ adipose progenitors. Cell Discov., 3, 17036.
95) Kong, J. & Li, Y.C. (2006) Molecular mechanism of 1,25-dihydroxyvitamin D3 inhibition of adipogenesis in 3T3-L1 cells. Am. J. Physiol. Endocrinol. Metab., 290, E916–E924.
96) Yao, Y., Zhu, L., He, L., Duan, Y., Liang, W., Nie, Z., Jin, Y., Wu, X., & Fang, Y. (2015) A meta-analysis of the relationship between vitamin D deficiency and obesity. Int. J. Clin. Exp. Med., 8, 14977–14984.
97) Jiang, H., Xiong, D.H., Guo, Y.F., Shen, H., Xiao, P., Yang, F., Chen, Y., Zhang, F., Recker, R.R., & Deng, H.W. (2007) Association analysis of vitamin D-binding protein gene polymorphisms with variations of obesity-related traits in Caucasian nuclear families. Int. J. Obes., 31, 1319–1324.
98) Ye, W.Z., Reis, A.F., Dubois-Laforgue, D., Bellanne-Chantelot, C., Timsit, J., & Velho, G. (2001) Vitamin D receptor gene polymorphisms are associated with obesity in type 2 diabetic subjects with early age of onset. Eur. J. Endocrinol., 145, 181–186.
99) Narvaez, C.J., Matthews, D., Broun, E., Chan, M., & Welsh, J. (2009) Lean phenotype and resistance to diet-induced obesity in vitamin D receptor knockout mice correlates with induction of uncoupling protein-1 in white adipose tissue. Endocrinology, 150, 651–661.
100) Wong, K.E., Szeto, F.L., Zhang, W., Ye, H., Kong, J., Zhang, Z., Sun, X.J., & Li, Y.C. (2009) Involvement of the vitamin D receptor in energy metabolism: regulation of uncoupling proteins. Am. J. Physiol. Endocrinol. Metab., 296, E820–E828.
101) Wong, K.E., Kong, J., Zhang, W., Szeto, F.L., Ye, H., Deb, D.K., Brady, M.J., & Li, Y.C. (2011) Targeted expression of human vitamin D receptor in adipocytes decreases energy expenditure and induces obesity in mice. J. Biol. Chem., 286, 33804–33810.
102) Malloy, P.J. & Feldman, B.J. (2013) Cell-autonomous regulation of brown fat identity gene UCP1 by unliganded vitamin D receptor. Mol. Endocrinol., 27, 1632–1642.
103) Schwartz, M.W., Peskind, E., Raskind, M., Boyko, E.J., & Porte, D. Jr. (1996) Cerebrospinal fluid leptin levels: relationship to plasma levels and to adiposity in humans. Nat. Med., 2, 589–593.
104) Cheng, A., Uetani, N., Simoncic, P.D., Chaubey, V.P., Lee-Loy, A., McGlade, C.J., Kennedy, B.P., & Tremblay, M.L. (2002) Attenuation of leptin action and regulation of obesity by protein tyrosine phosphatase 1B. Dev. Cell, 2, 497–503.
105) Bjorbak, C., Lavery, H.J., Bates, S.H., Olson, R.K., Davis, S.M., Flier, J.S., & Myers, M.G. Jr. (2000) SOCS3 mediates feedback inhibition of the leptin receptor via Tyr985. J. Biol. Chem., 275, 40649–40657.
106) Glick, Z. (1982) Inverse relationship between brown fat thermogenesis and meal size: the thermostatic control of food intake revisited. Physiol. Behav., 29, 1137–1140.
107) Okamatsu-Ogura, Y., Uozumi, A., Toda, C., Kimura, K., Yamashita, H., & Saito, M. (2007) Uncoupling protein 1 contributes to fat-reducing effect of leptin. Obes. Res. Clin. Pract., 1, 223–290.
108) Okamatsu-Ogura, Y., Nio-Kobayashi, J., Iwanaga, T., Terao, A., Kimura, K., & Saito, M. (2011) Possible involvement of uncoupling protein 1 in appetite control by leptin. Exp. Biol. Med. (Maywood), 236, 1274–1281.
109) Yamada, T., Katagiri, H., Ishigaki, Y., Ogihara, T., Imai, J., Uno, K., Hasegawa, Y., Gao, J., Ishihara, H., Niijima, A., et al. (2006) Signals from intra-abdominal fat modulate insulin and leptin sensitivity through different mechanisms: neuronal involvement in food-intake regulation. Cell Metab., 3, 223–229.