1)グルタミン
グルコースとグルタミンは,増殖する培養細胞の基本的な栄養因子であり,タンパク質,脂質および核酸の生合成に寄与している.両者は一般的な組織培養培地の中で最も豊富に含まれる添加物であり,解糖系・ペントースリン酸経路・TCA回路を介して異化され,ATPおよびNAD(P)を生成する.培養細胞でのタンパク質合成には培養液から直接取り込まれたアミノ酸と,グルコースおよびグルタミンから生合成された非必須アミノ酸とが利用される.さらに,グルコース由来のアセチルCoAは脂質合成に利用され,核酸合成においては,グルコースはリボースを提供し,グルタミンは窒素供給源として働く.PSCの活発な細胞増殖も外因性のグルコースとグルタミンに大きく依存しており,細胞増殖は両方が豊富なときに最大化される.PSCは,未分化なナイーブ型とやや分化が進んだプライム型に分類される.ナイーブ型とプライム型ではコロニー形態,遺伝子発現,未分化性維持シグナル,X染色体の活性化および代謝などに違いがある.グルタミンやその代謝物であるαKGはPSCで利用されるが,その作用はナイーブ型とプライム型との間で違いがある.マウスESC(mESC)などのナイーブ型PSCは特殊な代謝特性を持っており,グルコース由来の炭素からTCA回路を介してグルタミンを生成することが可能であり,グルタミン非存在下でも増殖することが可能である1).一方,ヒトESC(hESC)などのプライム型PSCでは,アコニターゼ2(aconitase 2:ACO2)およびイソクエン酸脱水素酵素2/3(isocitrate dehydrogenase 2/3:IDH2/3)の発現が低く,TCA回路においてピルビン酸由来のクエン酸を十分に利用することができないため,αKGを介したグルタミン生合成ができない2).そのため,プライム型PSCの生存にはグルコースとグルタミン両者が必須である.グルタミンによる未分化性維持機構についてはmESCを用いて,そのシグナル伝達経路が詳細に検討された.グルタミン除去により未分化マーカーであるOct4遺伝子や細胞周期関連遺伝子の発現が低下し,グルタミン添加はAkt, PKC(protein kinase C),mTOR(mammalian target of rapamycin)の活性化を引き起こし,mESCの増殖を刺激し,未分化性を維持していることが明らかとなった3).さらに,グルタミンはHDAC1(histone deacetylase 1)発現低下を誘導し,ヒストンアセチル化とメチル化の増加をもたらした.また,DNMT1(DNA methyltransferase 1)およびDNMT3a(DNA methyltransferase 3a)の発現減少を介して,Oct4プロモーター領域のDNAメチル化を顕著に低下させた.これらの結果からグルタミンはエピジェネティック修飾制御による未分化性維持に関与していることが示唆された.
体性幹細胞である造血幹細胞においてもグルコースとグルタミンは重要な栄養素である.造血幹細胞では取り込み型グルタミントランスポーター(ASCT2)が高発現しており,赤血球系への分化ではヌクレオチド生合成のためにグルタミンとグルコースが必要である4).そのため,グルタミンおよびグルコースの細胞内取り込みを阻害することによりヌクレオチド合成が障害されると,造血幹細胞の分化は赤血球系から骨髄系へと移行する.
また,グルタミンからグルタミン酸を経由して合成されるαKGはヒストン脱メチル化酵素(JmjC-domain containing histone demethylase:JHDM)やTET(ten-eleven translocation)といったエピジェネティック制御酵素を活性化させ,ヒストンおよびDNAの脱メチル化を誘導することで多能性関連遺伝子の発現を制御する.ナイーブ型PSCではαKG添加により自己複製が促進される一方で分化が抑制される1).mESCにおいて細胞内αKG濃度は,Oct4/Sox2/Nanogの標的タンパク質であるホスホセリンアミノトランスフェラーゼ1(phosphoserine aminotransferase 1:Psat1)によって調節される.Psat1の発現の低下は,DNAヒドロキシメチル化低下およびH3K9me3上昇を引き起こし,mESCの分化を加速させる5).一方,αKGはナイーブ型PSCの未分化性維持を担っているが,分化段階が若干進んだプライム型PSCにおいては,多能性遺伝子の発現を抑えることで分化を促進することが報告されている6).
このように,培養液中の最も基本的な栄養素であるグルタミンが未分化性維持や分化系譜決定において不可欠な役割を果たすことがわかり,アミノ酸による幹細胞の分化制御が重要視されることとなった.
2)トレオニン,グリシン,セリン
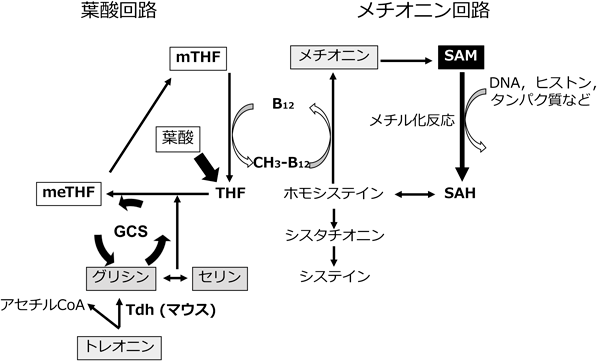
1炭素代謝はメチオニンと葉酸の代謝回路を含み,DNA,ポリアミン,クレアチン,リン脂質の合成にメチル基を提供するシステムであり,生体内でメチル基供与体として働くSAMもこの経路で生合成される7).1炭素代謝系にはトレオニン,グリシン,セリン,メチオニン,システインといったアミノ酸が含まれ,これらのアミノ酸はSAMを介して幹細胞でエピジェネティックな制御を行っている8).幹細胞におけるアミノ酸代謝研究の幕を開けたのは,2009年にScience誌に報告されたmESCのトレオニン依存性に関する論文である9).Wangらは,まずメタボローム解析を行い,マウス未分化ES細胞では1炭素代謝経路に炭素を供給するトレオニンが分化細胞と比較して極端に低いことを見いだした(図1).未分化細胞ではトレオニンからグリシンを合成する際の律速酵素であるトレオニン脱水素酵素(threonine dehydrogenase:Tdh),その下流のグリシンC-アセチルトランスフェラーゼ(glycine C-acetyltransferase:Gcat)とグリシンデカルボキシラーゼ(glycine decarboxylase:Gldc)は高発現しており,これがトレオニン濃度低下に寄与していた.低分子化合物を用いてTdhを阻害するか,または細胞培養液からトレオニンを完全に除去すると,増殖抑制,アポトーシス,および細胞周期の停止が引き起こされ,処理した細胞は死滅した9).その後の研究でトレオニンはAKT, MAPK,およびmTORをリン酸化することで,mESCの細胞増殖および未分化性維持に寄与することがわかった10).さらに,mESCをトレオニン除去培地で培養するとトレオニン由来のグリシンが低下し,葉酸回路を介して最終的にはメチオニンの代謝物でありメチル基供与体として働くSAMの細胞内濃度が低下して,ヒストンのメチル化修飾が低下することが明らかとされた11).これら一連のトレオニンについての研究によりアミノ酸が幹細胞においてシグナル伝達,およびエピジェネティック修飾に関与し,未分化性維持に働いていることが解明された.
1炭素代謝を構成するグリシンはトレオニンから生合成され,トレオニンによる多能性維持に中間代謝物として働くことがわかっていたが,それ自身による未分化性維持機構は十分に解明されていなかった.近年,グリシン開裂系(glycine cleavage system:GCS)酵素であるGldcの発現が幹細胞マーカーであるSox2とLin28Aによって制御されることが明らかになった.さらにGCSはグリシンからSAMの合成を増加させヒストンH3のリシン4トリメチル化(H3K4me3)修飾を促進するとともに,過剰なグリシンを分解し,細胞の老化を防ぐことで,PSCの未分化性を制御することが報告された12).
セリンとグリシンは可逆的に相互変換される関係にあり,1炭素代謝系を介して幹細胞のエピゲノム修飾に寄与する.Lin28A/Bのダブルノックアウト(KO)mESCを用いた詳細な代謝研究により,未分化細胞においてLIN28がグルコースからのセリン合成およびセリンからSAMへの代謝を正に制御することが報告され,未分化性維持へのセリンの寄与が再認識された13).さらに,酵母を用いた研究でセリン代謝酵素,SAM合成酵素およびヒストンリン酸化酵素が複合体を形成し,H3K4メチル化酵素と相互作用することが報告された14).この報告はSAMが特定のヒストンメチル化酵素において選択的に利用される可能性を示唆するもので興味深い.
3)メチオニン
マウスにおいてトレオニンはTdhによりグリシンとアセチルCoAに変換され,グリシンはH3K4me3を制御するために利用されるが,ヒトにおいては上記のTDHは発現しておらず,1炭素代謝の一部であるメチオニン回路自身がH3K4me3を制御していた15).我々は,まず必須アミノ酸9種類(トレオニン,バリン,イソロイシン,ロイシン,リシン,ヒスチジン,フェニルアラニン,トリプトファン,メチオニン)をそれぞれ一つずつ除去した培養液を作製し,hESC/ヒトiPSC(hiPSC)を2日培養した後の細胞数を計測した.その結果,ロイシン,リシン,トリプトファンおよびメチオニン除去により細胞数が有意に減少したが,メチオニン除去による効果が一番大きく,細胞数は約5%以下まで減少した.メチオニン除去はマウスにおけるトレオニン除去と同様に細胞内SAM濃度の低下を引き起こし,H3K4me3が顕著に低下し,p53タンパク質の発現増加,細胞周期停止に続いて,細胞死が起こることが確認された.さらにhESC/hiPSCの細胞分化において細胞死が起こらない程度の短時間(10時間)のメチオニン除去前処理を行うことで,NANOG発現低下が引き起こされ,その後の三胚葉への分化が起こりやすい遷移状態に変化した.興味深いことに,最近の研究ではSIRT1発現の下流でmESCの維持にもメチオニンが関与していることが示唆されている16).SIRT1 KO mESCは,メチオニンのSAMへの変換を触媒するメチオニンアデノシルトランスフェラーゼ2a(methionine adenosyltransferase 2a:MAT2a)の発現低下によりメチオニン/SAMが上昇する.メチオニン代謝の制御は,MAT2aプロモーターに結合してその発現を誘導するc-MYCおよびn-MYCのSIRT1依存性タンパク質発現を介して行われることも明らかにされた.さらに,マウスの小腸オルガノイドを用いた培養においても,培養液からのメチオニンの除去で未分化マーカーが減少し,分化を促進することが明らかにされ,応用が進んでいる17).
4)プロリン
プロリンは細胞外マトリックスであるコラーゲン中に大量に存在し,生体内において表皮細胞増殖促進,角質層保湿作用,コラーゲン合成促進活性を有するが,マウスPSC(mPSC)の分化促進効果も近年明らかにされてきた.循環母体血中および臍帯静脈血中にはプロリンが高濃度(~150 μM)に含まれており18),同程度の濃度のプロリンはmTORシグナル活性化を介してmESCの原始外胚葉への分化誘導を促進する19).さらに,プロリンはH3K9およびH3K36メチル化を増加させることで,エピゲノム制御にも関与しESCから間葉系細胞への分化を促進した20).
5)アルギニン
アルギニンは生体内生成速度が十分ではない成長期には摂取が必要なため準必須アミノ酸に分類されており,生体内では一酸化窒素の前駆体として働くとともにインスリンなどの分泌刺激,免疫反応の活性化,クレアチン生合成への利用など多様な作用を有する.幹細胞においても他のアミノ酸と相互作用することが報告されており,hESCの培養液からアルギニンとプロリンを同時に除去して48時間培養することで未分化マーカーであるNANOGおよびOCT4タンパク質の発現が顕著に低下し,胚様体分化では培養12日目において神経マーカーであるNESTINやTUJ1の発現が除去群で増加すること報告された21).プロリンは主に肝臓と小腸でグルタミン酸およびオルニチンからグルタミン酸セミアルデヒド,ピロリン-5-カルボン酸(pyrroline-5-carboxylate:PCA)を経て,生成される.さらに肝臓ではオルニチンは尿素回路でアルギニンから生合成される.一方,多能性幹細胞におけるプロリンとアルギニンの代謝相互作用については不明であり,詳細な検討が今後行われることを期待する.
また,アルギニンはメチオニン代謝との相互作用が報告されている22).PSCにおけるメチオニンの役割については前述したが,がん細胞においてもメチオニン除去はSAM低下を引き起こし,ヒストンメチル化修飾を低下させた.がん細胞ではSAMはヒストン修飾とともにクレアチン合成におけるメチル化反応にも多く利用されていた.生体内では,グリシンとアルギニンからグリコシアミンが合成され,これがグアニジノ酢酸N-メチルトランスフェラーゼによるメチル化反応によりエネルギー貯蔵物質であるクレアチンへと変換される.非常に興味深いことに上記のがん細胞においては,培養液からメチオニンとともにアルギニンを除去しておくことでクレアチン経路におけるSAM使用量が減少し,余剰となったSAMがヒストンメチル化に転用されることでH3K4me3が維持され,メチオニン除去特異的な遺伝子発現変化を抑制可能であった.
6)バリン,ロイシン
アミノ酸の代謝は主に肝臓で行われるが,分枝アミノ酸であるバリン,ロイシン,イソロイシンの代謝は筋肉,脂肪組織,脳などで行われる.これは分枝アミノ酸(分岐鎖アミノ酸)の異化反応酵素である分枝アミノトランスフェラーゼ(branched-chain aminotransferase:BCAT)が成熟肝臓には発現しないからである.一方,肝臓が発生初期に一過性増殖を示す事実を元にトランスクリプトーム解析およびメタボローム解析が行われ,マウスおよびヒト肝臓前駆細胞はBCATを時期特異的に発現しており,分枝アミノ酸を活発に利用できることが明らかにされた23).さらに,妊娠中の母親マウスへの分枝アミノ酸制限は,仔の肝臓前駆細胞の増殖を阻害した.分枝アミノ酸の中でも特にバリンの補充はヒトPSC(hPSC)由来肝臓オルガノイドのin vitro増殖を促進し,最終的にはバリンに富む特殊組成培地を用いた新たな分化誘導方法が構築された.これまでヒトにおけるin vitroアミノ酸代謝研究は,がん細胞や最終分化した初代培養細胞を用いて行われてきたが,今後はhPSC分化誘導系を利用することにより発生分化段階に沿った検討が可能となり,今回の胎生期肝臓における分枝アミノ酸の利用のような新たな知見が得られることが期待される.
このバリンについては非常に興味深い知見が報告された.マウスおよびヒトの造血幹細胞は他のアミノ酸と比較して特にバリン要求性が高く,マウスにバリン制限食を与えると骨髄の造血幹細胞の減少が引き起こされ,その後の造血幹細胞移植が成立した24).このバリン制限食を用いた前処置は,従来の抗がん剤や全身放射線照射による処置と比較して毒性を大幅に低減できる手法として注目を集めている.
分枝アミノ酸の一つであるロイシンは体組織と食物タンパク質に最も多く,1日の必要量がアミノ酸の中で最大である.さらにロイシンはmTORを活性化し,タンパク質合成を促進するとともに分解を抑制する.ロイシンは幼児には成長に,成人でも窒素平衡の維持に必須であるが,mTOR活性化を介して胎生期の膵臓発生分化に影響を与えることが報告された25).妊娠ラットに食事性ロイシンを補給すると,仔マウスは膵臓β細胞量が減少し,耐糖能が低下した.胎仔膵臓におけるmTOR活性化は,転写因子Ngn3の抑制因子として機能する低酸素誘導因子(hypoxia inducible factor:HIF)の発現増加を導くことから,過剰な食事性ロイシンによるmTOR活性化は,Ngn3陽性内分泌前駆細胞を減少させ,そこから分化する成熟β細胞の数が減少したと考えられる.
7)トリプトファン(セロトニン)
必須アミノ酸であるトリプトファンは生体内において二つの重要な代謝経路で分解される.一つは,インドールアミン2,3-ジオキシゲナーゼ1(indoleamine 2,3-dioxygenase 1:IDO1)の反応でインドールが開裂してキヌレニンを合成する経路で,ニコチン酸,ニコチンアミド,NAD(P)が合成される.未分化hPSCは分化細胞と比較してIDO1活性が非常に高いため,キヌレニンが多く産生されており,アリール炭化水素受容体(aryl hydrocarbon receptor:AhR)を刺激して自己複製遺伝子の発現を促進することが明らかにされた.また,PSCを分化させると,キヌレニンアミノトランスフェラーゼを中心としたキヌレニン異化経路が活性化され,培地中のキヌレニンの量が減少し,分解産物である2-アミノアジピン酸(2-aminoadipic acid:2-AAA)が培地中に分泌された.このようにトリプトファン由来のキヌレニンの代謝は,AhRを介して未分化性維持に重要な役割を果たしており,培養液中に分泌されたキヌレニンおよび2-AAAはそれぞれ未分化細胞および分化細胞のマーカーとして利用可能であることが示された26).
トリプトファンのもう一つの代謝経路は,脱炭酸反応により生理活性アミンのセロトニンを生じるものである.セロトニンは神経伝達物質として働くとともに,ニューロンの生存,および分化を助ける栄養因子として機能し,妊娠期においては膵臓β細胞の複製やグルコース応答性を制御する27).また,セトロニンはタンパク質に結合するセロトニン化といわれる翻訳後修飾により,細胞のシグナル伝達を変化させることができる.近年,核タンパク質であるヒストンH3のセロトニン化修飾が,神経細胞の分化を抑制することが報告され,栄養因子由来の代謝物によるクロマチン修飾を介した新たな分化制御機構として関心を集めている28).
8)ヒスチジン(ヒスタミン)
ヒスチジンは生体アミンとしても知られるヒスタミンの起源物質である.ヒスタミンはL-ヒスチジン脱炭酸酵素(L-histidine decarboxylase:HDC)による脱炭酸反応により生合成され,ヒスタミンH1受容体を介して平滑筋収縮やアレルギー反応を引き起こす.HDC KOマウスにおけるヒスタミン欠乏症はマクロファージの浸潤障害と心筋細胞のアポトーシスを介して急性心筋梗塞の心筋損傷を悪化させる29, 30).最近の研究では,HDCは造血幹細胞や骨髄前駆細胞に発現しており,生合成されたヒスタミンはH2受容体を介して自己複製を促進することが明らかになった31).また,ヒスタミンはH1受容体を介してマウス神経幹細胞の分化を促進する32).さらに,ヒスタミンはERK1/2-STAT3シグナル伝達経路を介してhiPSCからの心筋分化を有意に促進することが報告された33).これらのデータは,ヒスチジン由来の生理活性アミンであるヒスタミンが幹細胞の発生と分化に重要な役割を果たしていることを示唆している.
9)フェニルアラニン,チロシン(ドパミン)
ヒトは生体内でベンゼン環を生合成できないので,フェニルアラニンを必須アミノ酸として食事から摂取している.フェニルアラニンからはチロシンを経て,多くの生理活性アミンが合成される.我々はmESC由来の膵臓前駆細胞から内分泌前駆細胞への分化を促進する薬剤をスクリーニングした結果,小胞モノアミン輸送体の一つであるVMAT2(vesicular monoamine transporter 2)の阻害剤であるレセルピンおよびテトラベナジンを同定した.さらにVMAT2により小胞への取り込みが制御されるドパミン,ヒスタミン,セロトニンなどモノアミンは,膵臓β細胞への分化を抑制することも見いだした34).さらに,マウス膵島を用いた実験でドパミンD2受容体(dopamine D2 receptor:DRD2)アンタゴニストであるドンペリドンがβ細胞数を増加させ,ヒト膵島においてドパミンがDRD2を介して膵臓β細胞量を制御することを見いだした35).このことから,フェニルアラニン,チロシン由来の生理活性アミンであるドパミンが分化制御因子として働くことが明らかとなった.
これまで述べたように,幹細胞の未分化性維持において,特殊なアミノ酸代謝機構が利用されている.そこで,この未分化細胞と分化細胞の代謝特性の違いを利用して目的細胞のみを選抜する種々の培養方法が開発されてきた.特に,非常に発達したミトコンドリアを持つ心筋や,アミノ酸を含め栄養代謝の本体である肝臓への分化において研究が進んでいる.この方法は,遺伝子改変や機械的な選抜作業を伴わず,特殊組成培地のみを利用するため,簡便であり大量培養への応用へ向いている.以下にこれまで報告された選抜培養方法を紹介するとともに,そこで利用されている代謝特性について議論したい.
胎児の心臓は,酸化的リン酸化を介してATP合成を行う際にグルコースよりも乳酸を基質として利用することが知られていた36).これらの知見や遺伝子発現および代謝解析結果から,乳酸を含むグルコース不含培地がPSCs由来の心筋細胞の選抜に有効であると仮説が立てられ,hESCまたはmESCから心筋分化した胚様体をグルコース不含乳酸培地で培養することで心筋細胞を95%以上に濃縮することに成功した37).さらに,同グループはhESC/iPSCにおけるグルコースおよびグルタミン代謝の役割を明らかにし,乳酸を含むグルコース不含培地からさらにグルタミンを除去し,心筋分化誘導後の選択培地として利用した.その結果,腫瘍形成の原因となる未分化幹細胞を死滅させ,心筋細胞だけを生きたまま選別する新たな方法を確立した2).
肝臓は薬物代謝の中心部位であり,ここでは複数の肝臓特異的な代謝経路が作用している.これらの肝臓に存在する複数の代謝経路を利用して,mESCおよびhiPSC由来肝臓細胞の選択的培地が開発された38, 39).肝臓では尿素回路によりオルニチンからアルギニンが生合成され,フェニルアラニン水酸化酵素によってフェニルアラニンからチロシンが作られる.さらにピルビン酸キナーゼによってホスホエノールピルビン酸からピルビン酸が生成され,グリセロールとガラクトースから,グリセロールキナーゼとガラクトキナーゼを介してグルコースが合成される.これらの肝臓が持つ代謝系を考慮して,選択培地としてはアルギニン,チロシン,ピルビン酸,およびグルコース不含で,それらの基質となるオルニチン,フェニルアラニン,ガラクトース,およびグリセロールが豊富な培地が開発され,肝臓細胞の濃縮に利用可能であることが証明された.より直接的な方法として肝臓が持つ高いアンモニア処理能に着目して,分化細胞にアンモニアを添加することでアンモニアの毒性に耐えられない未分化細胞や非肝臓系の細胞を死滅させ,肝臓細胞を選抜する方法も報告された40).
未分化細胞では,メチオニン代謝回路に関わる酵素群の多くが高発現し,内胚葉に比べてメチオニン消費が約5倍高い.そこで,内胚葉分化期間の後半2日間をメチオニン除去した分化誘導培地に替えると,未分化細胞選択的に細胞死が誘導され,相対的に内胚葉の割合が増加し,その後の肝臓の分化効率も増加した15).分化過程でメチオニン除去培地を利用することで残存する未分化細胞の除去が可能であり,分化効率が悪い細胞株を利用する場合には非常に有効である.また,メチオニン除去培地はhiPSCからの心筋細胞分化においても未分化細胞除去に利用可能である41).