マクロピノサイトーシスを活用した細胞内送達の可能性Potential of macropinocytosis for intracellular delivery
京都大学化学研究所Institute for Chemical Research, Kyoto University ◇ 〒611–0011 京都府宇治市五ヶ庄 ◇ Gokasho, Uji, Kyoto 611–0011, Japan
発行日:2021年2月25日Published: February 25, 2021
© 2021 公益社団法人日本生化学会© 2021 The Japanese Biochemical Society
近年,siRNAやアンチセンス核酸,抗体などのタンパク質,活性構造を化学的に増強したペプチドなどをはじめとした,種々のバイオ医薬品の開発が盛んに行われるようになった.これらの中には血中や細胞表面の分子と相互作用し,薬効を示すものもあるが,細胞内に移行して初めて効果を示すものも多い.分子量が500程度までで適度な親水性と疎水性のバランスを有する小分子薬物の中には,受動拡散により細胞膜を透過し,細胞内に移行するものも多い.しかし,上記のようなバイオ医薬品は分子量が大きい上に一般に親水性が高く,細胞膜にほとんど浸透しない.このような分子を細胞内に送達する際には,エンドサイトーシスによりいったん細胞内に取り込ませ,次いで,何らかの手段でエンドソームからサイトゾルに移行(エンドソーム脱出)させるアプローチが一般にとられる.エンドソーム脱出を促進させる作用を持つ高分子やリポソーム,ペプチド等の開発も盛んに行われている1).しかし,目的物質がエンドソーム内に移行することなしには,これらの系は効果を発揮しえない.つまり,エンドソーム脱出を促進させる系を活用するには,これに先立ちバイオ医薬品の細胞外からエンドソーム内への移行量を高めることが重要である.細胞外分子をエンドソームに効果的に封入する細胞機能として,受容体依存性エンドサイトーシスがある.リガンドと細胞表面の受容体との相互作用により,リガンドは効果的にエンドソーム内に封入される.しかし,細胞内に取り込ませたいバイオ医薬品に対して,標的細胞が適当な受容体を持っているとは限らない.
筆者らは,タンパク質・ペプチドの細胞内送達に有用なアルギニンに富む塩基性ペプチド(HIV-1 TATペプチドやオリゴアルギニンなど:以下,アルギニンペプチドと記載)の細胞内移行にマクロピノサイトーシスと呼ばれるアクチン駆動型の液相エンドサイトーシスが重要な役割を果たすことを報告している2).一方では,ここ数年の研究により,マクロピノサイトーシスには,従来考えられていた以上にさまざまな様式のものがあることが見いだされ(表1),がん細胞の養分摂取に重要な役割を果たす可能性も示唆されている3).本稿では,マクロピノサイトーシスの理解の現況を概説するとともに,細胞内へのバイオ医薬品の送達におけるマクロピノサイトーシスの可能性に関して,筆者らの知見を併せ紹介したい.
| 刺激誘導型 |
| 内因性 |
| 成長因子(EGF, PDGFなど) |
| ケモカイン(SDF-1α) |
| 分泌タンパク質(Wnt-3a) |
| 外因性 |
| ウイルス(アデノウイルスなど) |
| ホルボールエステル(phorbol myristate acetate:PMA) |
| リポ多糖(LPS) |
| 常時活性型 |
| マクロファージ[カルシウム感知受容体(CaSR)依存的] |
| Ras変異がん細胞(KRAS G12V変異など) |
代表的なエンドサイトーシスとしてクラスリン依存性エンドサイトーシスやカベオラ依存性エンドサイトーシスが知られる.これらにおいては,細胞膜が積荷分子とともに細胞内に陥没し,クラスリンやカベオリンによる膜の被覆化を伴って膜小胞が形成される.一方,マクロピノサイトーシスでは,外的刺激に呼応して細胞骨格タンパク質のアクチンの重合により,細胞外に向けて細胞膜が突出後,細胞膜との融合が起こり,巨大なサイズの膜小胞(マクロピノソーム)が形成される4).外的刺激としてはEGF, PDGFなどの増殖因子,ストロマ細胞由来因子1α(SDF-1α)などのケモカイン,あるいはアデノウイルス等が知られている.Wnt-3aによる刺激によりマクロピノサイトーシスが誘導されることも最近報告された(図1)5).
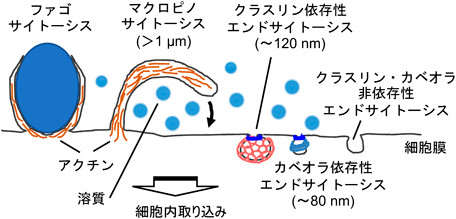
クラスリン依存性エンドサイトーシスやカベオラ依存性エンドサイトーシスにより生じる小胞の直径がそれぞれ120 nm, 80 nm程度であるとされるのに対し,マクロピノソームの直径は1 µm~数µmにも達することが報告されている6).そのため,マクロピノソーム形成時に細胞外液とその溶質分子,あるいは細胞表面に付着していた細胞外分子が非特異的にマクロピノソーム内に取り込まれると考えられている.顕微鏡観察においては,クラスリン依存性・カベオラ依存性エンドサイトーシスや,小胞サイズの小さいクラスリン・カベオラ非依存性エンドサイトーシスと区別するために,これらのエンドサイトーシスではほとんど取り込まれないサイズの大きな溶質として70 kDaデキストランが一般的なマクロピノソームマーカーとして使用される5, 7).
一方,マクロファージなどの貪食系の細胞では,リポ多糖(LPS)刺激などでマクロピノサイトーシスが誘導されることも知られているが,恒常的にもマクロピノサイトーシスが起こっており,生体防御の一端を担うとされている8).この恒常的なマクロピノサイトーシス活性化には,カルシウム感知受容体(calcium sensing receptor:CaSR)が関与することが示唆されている.非マクロファージ系の細胞として,Ras変異を有するがん細胞においても,マクロピノサイトーシスが外的刺激によらず恒常的に起きていることが報告されている9, 10).これらのがん細胞では,マクロピノサイトーシスにより血清タンパク質(アルブミンなど)を積極的に細胞内に取り込み,エネルギー源としていることが報告され,非がん細胞における取り込み量との差を利用したがん治療への応用の可能性に関しても言及されている.マクロファージなどでは,古くなった赤血球細胞や細菌等の貪食に関わるファゴサイトーシスと呼ばれるエンドサイトーシスも行われている.アクチン駆動性である点でマクロピノサイトーシスと共通するが,ファゴサイトーシスは,(i)マクロファージ等の貪食細胞において,細菌などの病原微生物や死細胞などの大きな異物を細胞内に取り込むエンドサイトーシス(食作用)の一様式であり,(ii)取り込む異物の周囲に細胞膜を密着させて,細胞内への取り込みを行う.これに対し,マクロピノサイトーシスは貪食細胞以外でもみられる細胞外液を取り込むエンドサイトーシス(飲作用)であり,取り込む物質と細胞膜の密着は特に必要ないことで区別される4).
外因性物質の細胞内送達という観点からは,マクロピノサイトーシスは他のエンドサイトーシスとは二つの点で異なる特徴を有すると筆者らは考えている.一つはその取り込みサイズである.上述のようにクラスリン,あるいはカベオラ依存性のエンドサイトーシスの小胞の直径は100 nm前後であるとされるのに対し,マクロピノサイトーシスでは1 µmを超える小胞が形成されるとされている.直径数十nmあるいはそれ以上のナノ粒子が細胞内に移行しようとするとき,物理的にクラスリンエンドサイトーシスやカベオラエンドサイトーシスにより形成される小胞には入り込みにくいことが容易に予想される.
もう一つはマクロピノサイトーシスにおける取り込みの非特異性である.クラスリンエンドサイトーシスは,一面ではクラスリン被覆ピットとアダプター分子を介して相互作用可能な受容体を用いて,細胞の営みに必要な特定の養分等を,積極的にエンドソーム内に取り込む経路と捉えることができる.一方,マクロピノサイトーシスは細胞表面分子と相互作用する粒子や細胞付近に存在する溶質を一網打尽に細胞内に取り込む経路と考えられる.必ずしも特異的な相互作用は必要ない.したがって,標的細胞に目的物質を送達するために利用できる適切な受容体が存在しない場合でも,マクロピノサイトーシスを利用することで細胞内送達が促進されることが期待される.
例として細胞透過ペプチド(cell-penetrating peptide:CPP)を用いた細胞内送達があげられる.CPPとしては,HIV-1 TATペプチドやオリゴアルギニンなどのアルギニンペプチドが汎用されており,これらのペプチドとのコンジュゲーションによりさまざまなタンパク質や薬物が細胞内に効率的に送達され,所望の活性が得られることが報告されている2).これを達成できる理由の一つとして,アルギニンペプチドの細胞表面のヘパラン硫酸プロテオグリカン(HSPG)等との相互作用による細胞表面への濃縮とマクロピノサイトーシスの誘起による細胞内取り込みの促進が考えられている.アルギニンペプチドの他にも,炭化水素鎖で構造規制されたヘリックスペプチド(ステープルペプチド),種々のナノ粒子,ウイルス,細胞外小胞などもマクロピノサイトーシスで取り込まれる可能性が指摘されており11, 12),これまで詳細に調べられていなかっただけで,種々の外因性分子の細胞内移行にマクロピノサイトーシスが関連している可能性がある.
マクロピノサイトーシスを含むエンドサイトーシスでバイオ医薬品が細胞内に取り込まれたとしても,多くの場合はこれらの分子はリソソームなどに運搬されて分解される.したがって,細胞内送達という観点からは,エンドソーム内に保持されたバイオ医薬品が分解を受ける前に,エンドソームからサイトゾルに放出されることが必要と考えられる.エンドソーム膜の傷害により,これは可能となる一方,細胞膜も傷害すれば細胞に大きなダメージを与える.このため,エンドソーム膜を優先的に撹乱する方法論の開発がなされてきた.例として,エンドソームの成熟に伴うエンドソーム内のpH低下を利用して,エンドソーム膜を優先的に傷害するpH感受性ペプチドや高分子があげられる.一方,エンドソームの成熟化に伴い,構成脂質に変化が起こることも知られており,たとえば,後期エンドソームやリソソームの膜にはbis(monoacylglycero)phosphate(BMP)が多量に含まれることが示唆されている13).生体内での脂質のほとんどは中性もしくは酸性であることを考えると,BMPが多量に含まれる後期エンドソームやリソソーム膜も酸性を帯びていることが想定される.それならば,塩基性のペプチドや高分子は,中性や酸性のペプチドや高分子と比べて,後期エンドソームやリソソームの膜とより強く相互作用し,損傷することが可能かもしれない.一方,アルギニンペプチドは細胞表面への集積とマクロピノサイトーシスの誘起を通して,細胞内にバイオ高分子に送達する能力を持っているが,アルギニンペプチドのエンドソーム膜損傷能は実際にはそれほど高くなく,送達分子のエンドソーム内からのサイトゾルへの放出はたかだか数%程度と考えられている.また,送達分子の分子量やサイズが大きくなるに従い,その効率は悪くなることもいわれている.そのため,マクロピノサイトーシスの誘起による取り込み促進とエンドソームからサイトゾルへの送達分子の効果的な放出を可能とする,バイオ医薬品の細胞内送達における新たな方法論が求められていた(図2).
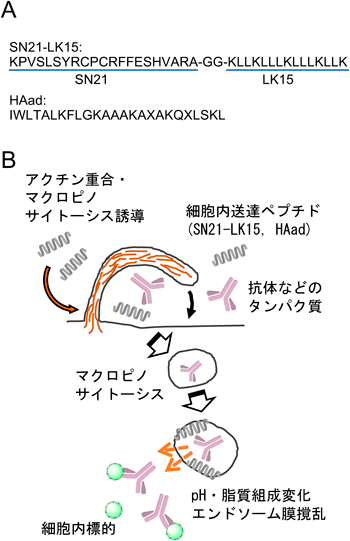
(A)細胞内送達ペプチドSN21-LK15とHAad[ペプチドのC末端はアミド;X=L-2-aminoadipic acid (Aad)],(B)マクロピノサイトーシスの誘起による取り込み促進とエンドソーム膜撹乱による送達分子のサイトゾルへの放出
我々の研究室では最近,SN21-LK15ペプチド(KPVSLSYRCPCRFFESHVARAGGKLLKLLLKLLLKLLK-amide)を開発した14).SN21ペプチドはSDF-1αのN末端21アミノ酸配列に対応するペプチドであり,高いマクロピノサイトーシス誘導能を持つ.LK15は塩基性のLysと疎水性のLeuからなる両親媒性ペプチドであり,酸性脂質に富む細菌の細胞膜を傷害する抗菌ペプチドとしても知られる.この二つのペプチドを連結させたSN21-LK15との共投与によりプラスミド,siRNA,抗体や種々のタンパク質が細胞内に送達され,それらの生物活性発現につながることが確認できた.
また,抗菌ペプチドとして見いだされたM-lycotoxinという塩基性ペプチドの配列に見直しを加えた細胞内送達ペプチドHAad[IWLTALKFLGKAAAKAXAKQXLSKL-amide;X=L-2-aminoadipic acid(Aad)]を最近発表した15).M-lycotoxinの抗菌性発揮には,分子中の疎水性アミノ酸と膜との相互作用による膜構造の撹乱が重要であると考えられる.HAadでは,(i)M-lycotoxin中の二つのヒスチジン残基がアラニンに置換されることで両親媒性ヘリックス構造の増強が図られ,また,(ii)その際形成される疎水面に,二つのAadが配置されている.Aadの側鎖カルボキシ基は細胞外では解離し,負電荷を有することで細胞膜との相互作用と細胞膜への傷害性が軽減される一方,分子全体としては塩基性を有しており,細胞表面に吸着するとともに,HAadならびに細胞内への送達分子のエンドソーム内への取り込みを促進する.エンドソーム内では,pH低下に伴い,電荷を持たない分子型(非解離型)が増えるために,ペプチドの疎水性が向上するとともに,ペプチドの塩基性によりエンドソーム膜との親和性が高まり,効果的にエンドソーム膜を破壊することが期待される.実際に,このペプチドとともに細胞に共投与することで,抗体などの大きなタンパク質の効果的な細胞内(サイトゾル)移送と細胞内標的分子との結合が確認された.
マクロピノサイトーシスは,これまで非特異的なピノサイトーシス経路として漠然と捉えられてきたが,種々のウイルスの感染やがん細胞の栄養補給経路としての役割が指摘されるに従い,ここ数年でいっそうの関心が集まっている.また,がん細胞への薬物の選択的送達の可能性も含めて,この経路の利用に関しても関心が高まっている.現時点では,非クラスリン依存性・非カベオラ依存性でサイズの大きなエンドソームを形成するピノサイトーシス=マクロピノサイトーシスという大雑把な理解しかされておらず,追跡マーカーとして70 kDaポリデキストランなどが用いられるだけである.マクロピノサイトーシスの誘導・調節機序や細胞内シグナル伝達との関連に関しても不明な点も多い.しかし,本稿で述べたように,マクロピノサイトーシスは実際には種々の機構で起こりうるものであり,細胞の営みや生体防御にこれまで考えられていた以上に重要な役割を果たす経路であるかもしれない.今後の研究の進展により,その理解と活用が進むことを期待したい.
1) Brock, D.J., Kondow-McConaghy, H.M., Hager, E.C., & Pellois, J.P. (2019) Endosomal Escape and Cytosolic Penetration of Macromolecules Mediated by Synthetic Delivery Agents. Bioconjug. Chem., 30, 293–304.
2) Futaki, S. & Nakase, I. (2017) Cell-Surface Interactions on Arginine-Rich Cell-Penetrating Peptides Allow for Multiplex Modes of Internalization. Acc. Chem. Res., 50, 2449–2456.
3) Zhang, Y. & Commisso, C. (2019) Macropinocytosis in Cancer: A Complex Signaling Network. Trends Cancer, 5, 332–334.
4) Mercer, J. & Helenius, A. (2009) Virus entry by macropinocytosis. Nat. Cell Biol., 11, 510–520.
5) Tejeda-Muñoz, N., Albrecht, L.V., Bui, M.H., & De Robertis, E.M. (2019) Wnt canonical pathway activates macropinocytosis and lysosomal degradation of extracellular proteins. Proc. Natl. Acad. Sci. USA, 116, 10402–10411.
6) Conner, S.D. & Schmid, S.L. (2003) Regulated portals of entry into the cell. Nature, 422, 37–44.
7) Commisso, C., Flinn, R.J., & Bar-Sagi, D. (2014) Determining the macropinocytic index of cells through a quantitative image-based assay. Nat. Protoc., 9, 182–192.
8) Doodnauth, S.A., Grinstein, S., & Maxson, M.E. (2019) Constitutive and stimulated macropinocytosis in macrophages: roles in immunity and in the pathogenesis of atherosclerosis. Philos. Trans. R. Soc. Lond. B Biol. Sci., 374, 20180147.
9) Ramirez, C., Hauser, A.D., Vucic, E.A., & Bar-Sagi, D. (2019) Plasma membrane V-ATPase controls oncogenic RAS-induced macropinocytosis. Nature, 576, 477–481.
10) Commisso, C., Davidson, S.M., Soydaner-Azeloglu, R.G., Parker, S.J., Kamphorst, J.J., Hackett, S., Grabocka, E., Nofal, M., Drebin, J.A., Thompson, C.B., et al. (2013) Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature, 497, 633–637.
11) Sakagami, K., Masuda, T., Kawano, K., & Futaki, S. (2018) Importance of Net Hydrophobicity in the Cellular Uptake of All-Hydrocarbon Stapled Peptides. Mol. Pharm., 15, 1332–1340.
12) Schneider, D.J., Speth, J.M., Penke, L.R., Wettlaufer, S.H., Swanson, J.A., & Peters-Golden, M. (2017) Mechanisms and modulation of microvesicle uptake in a model of alveolar cell communication. J. Biol. Chem., 292, 20897–20910.
13) Matsuo, H., Chevallier, J., Mayran, N., Le Blanc, I., Ferguson, C., Fauré, J., Blanc, N.S., Matile, S., Dubochet, J., Sadoul, R., et al. (2004) Role of LBPA and Alix in multivesicular liposome formation and endosome organization. Science, 303, 531–534.
14) Arafiles, J.V.V., Hirose, H., Akishiba, M., Tsuji, S., Imanishi, M., & Futaki, S. (2020) Stimulating Macropinocytosis for Intracellular Nucleic Acid and Protein Delivery: A Combined Strategy with Membrane-Lytic Peptides To Facilitate Endosomal Escape. Bioconjug. Chem., 31, 547–553.
15) Sakamoto, K., Akishiba, M., Iwata, T., Murata, K., Mizuno, S., Kawano, K., Imanishi, M., Sugiyama, F., & Futaki, S. (2020) Optimizing charge switching in membrane lytic peptides for endosomal release of biomacromolecules. Angew. Chem. Int. Ed. Engl., 59, 19990–19998.

1983年京都大学薬学部卒業,87年同大学院博士後期課程中退,徳島大学薬学部助手,93年同助教授,97年京都大学化学研究所助教授,2005年より現職.この間,1989年~91年米国ロックフェラー大学博士研究員.
研究テーマと抱負ペプチド・タンパク質の機能設計.特に生体膜と相互作用するペプチドの創出と,相互作用により得られる膜の構造や形態変化の解析と応用に興味を持って研究を進めています.
ウェブサイトhttps://www.scl.kyoto-u.ac.jp/~bfdc/index.html
趣味読書?

京都大学化学研究所特定准教授.博士(薬学).
1983年富山県魚津市に生る.2007年京都大学薬学部卒業.12年同大学院薬学研究科博士後期課程修了.フランスCNRSおよび東京大学理学系研究科でのポスドクを経て,19年より現職.
研究テーマと抱負エクソソームをはじめとする細胞外微粒子の細胞内取り込み機構と細胞内運命について.顕微鏡による可視化を軸にして生命現象の理解につながる研究をしていきたい.
趣味ジョギング.

This page was created on 2021-01-04T16:09:35.517+09:00
This page was last modified on 2021-02-04T16:14:59.000+09:00
このサイトは(株)国際文献社によって運用されています。