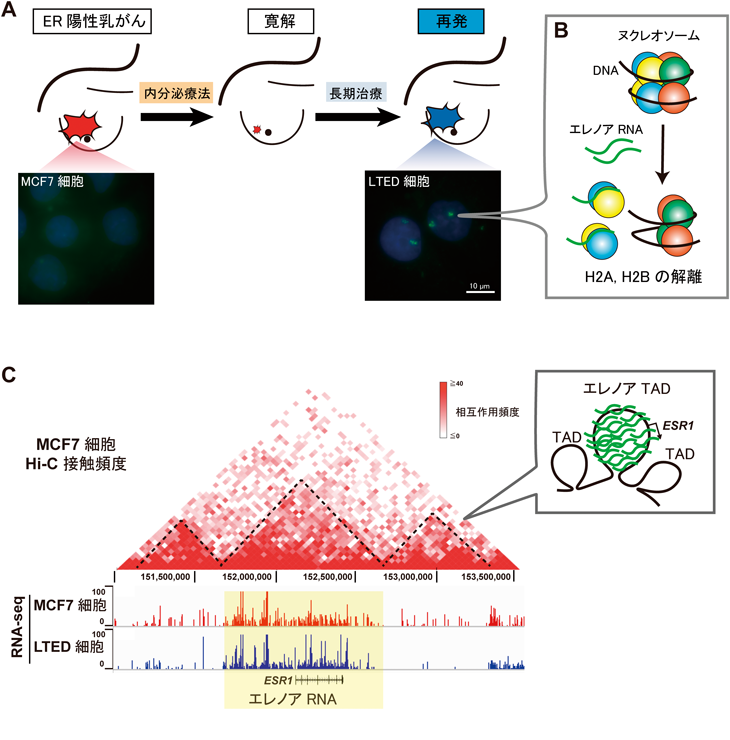
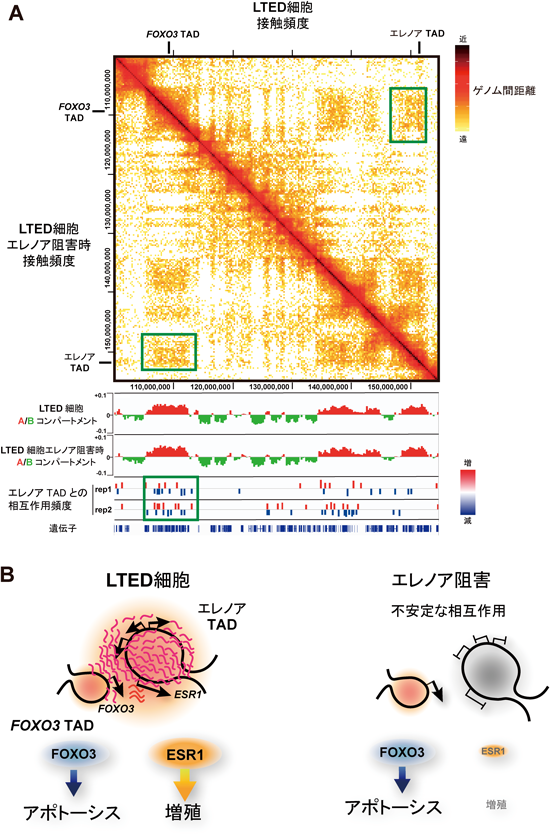
ノンコーディングRNAによる3次元ゲノム構造の制御Regulation of 3D genome structure by non-coding RNAs
がん研究会がん研究所がん生物部Division of Cancer Biology, The Cancer Institute of JFCR ◇ 〒135–8550 東京都江東区有明3–8–31 ◇ 3–8–31 Ariake, Koto-ku, Tokyo 135–8550, Japan
発行日:2021年2月25日Published: February 25, 2021