1. 長期増強現象とCa2+/カルモジュリン依存性タンパク質キナーゼ
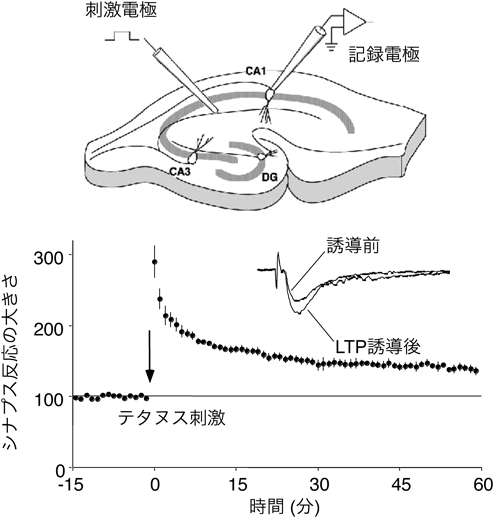
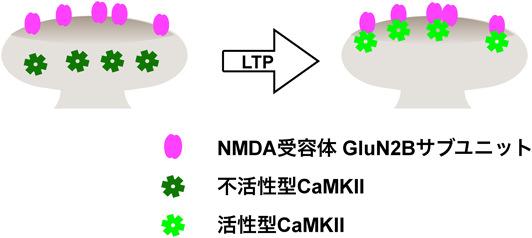
記憶形成時に同時に発火した神経細胞どうしのシナプスは,長期的に強化される.この性質をシナプス反応の長期増強現象[long-term potentiation(LTP),図1]と呼び,それにより,それらの細胞はその後も同時に発火する,「神経細胞集成体」を形成すると考えられる1).記憶想起時にはこの神経細胞集成体が再活性化されると考えられている.シナプス反応のLTPは1973年に海馬歯状回で報告されて以来2),記憶の細胞基盤として多くの興味を集めてきた.
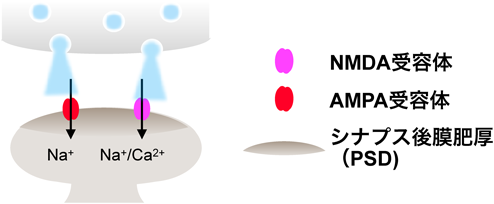
シナプスにはAMPA型グルタミン酸受容体(以下AMPA受容体)とNMDA型グルタミン酸受容体(以下NMDA受容体)の2種類のグルタミン酸受容体が存在する(図2).AMPA受容体は通常のシナプス伝達に寄与する一方,頻回刺激(記憶形成時の神経活動の亢進を模倣していると考えられる)により一過性にNMDA受容体が活性化し,カルシウムイオン(Ca2+)が細胞内に流入することがLTPを誘導する3).細胞内に流入したCa2+は細胞内伝達系を活性化し,その結果,最終的にAMPA受容体を介したシナプス伝達を増強する.Ca2+はタンパク質キナーゼCや一部のアデニル酸シクラーゼを活性化する他,カルモジュリンと結合し,Ca2+/カルモジュリン依存性タンパク質キナーゼを活性化させる.なかでも注目を集めてきたのは,タンパク質セリン/トレオニンキナーゼであるCa2+/カルモジュリン依存性タンパク質キナーゼII(CaMKII)である4).
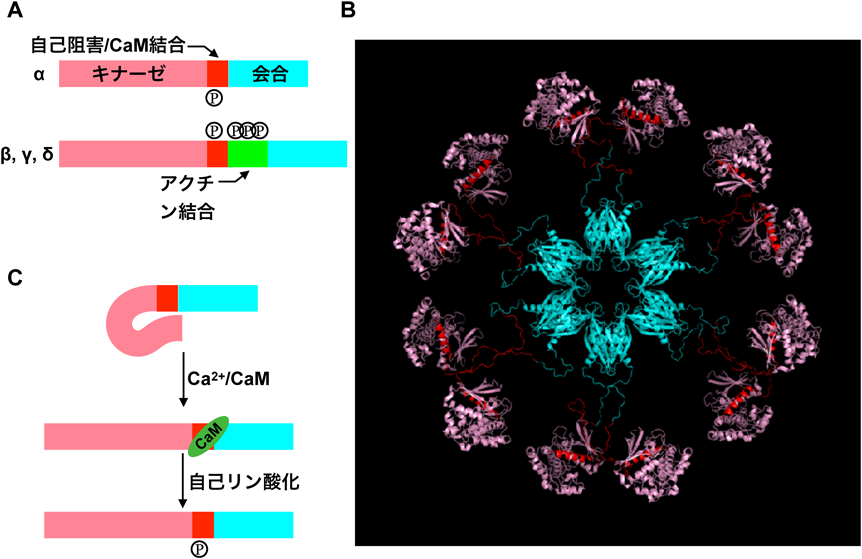
CaMKIIはセロトニン合成の律速段階酵素であるトリプトファン水酸化酵素の活性化因子として藤澤仁,山内卓ら5–7)によって,またほぼ同時期にその他いくつかのグループ8–11)によっても独立に発見された.CAMK2A(αサブユニット),CAMK2B(βサブユニット),CAMK2G(γサブユニット),CAMK2D(δサブユニット)の四つの遺伝子でコードされ(図3),それぞれ選択的スプライシングにより多数のバリアントを形成する12–14).
特筆すべきは,CaMKIIはCa2+/カルモジュリンにより活性化されると,自己阻害ドメインにあるトレオニン286(T286)が自己リン酸化されることで,自己阻害がかからなくなり,活性持続型になる点である15).そのことから,数秒の単位と考えられているLTP後のCa2+の上昇16)を,持続的なキナーゼ活性と変換させる機能があると予想され,これに基づきLismanらはCaMKIIに記憶分子としての役割を提唱した17).実際にCaMKIIの阻害薬を神経細胞に作用させるとLTPが阻害される18, 19).活性化CaMKIIの細胞内導入により,LTP様のシナプス反応が亢進し,かつそれ以上のLTPも起こらなくなった20–23).一方,CaMKII遺伝子を破壊した動物個体ではLTPのみならず,記憶学習も障害される24, 25).さらにT286をアラニン(A)に置換したノックイン動物も作製され,同様に記憶が障害されることも確認された26).これらの事実から,CaMKIIがLTPに密接に関与していることは明らかである.
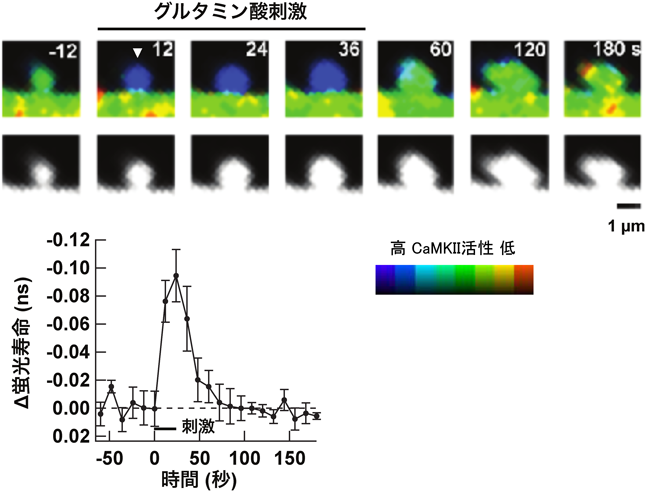
それでは,CaMKIIの活性化はLTP誘導後どの程度続くのであろうか.Kennedyらは抗リン酸化T286に対する抗体を作製し,LTP誘導によってT286のリン酸化が上昇するかを検討した27).その結果,LTP誘導後30分の時点で,T286リン酸化CaMKIIの量が上昇していることがわかった.福永らはスライス標品でLTPを誘導し,その後生化学的な解析をすることで,LTP後持続活性型のCaMKIIが上昇することを見いだしている28, 29).しかし一方,CaMKIIのFörster共鳴エネルギー移動(Förster Resonance Energy Transfer:FRET)センサーCamui30, 31)を用い,ケージ化グルタミン酸の脱ケージ化(グルタミン酸を光分解性の保護基により不活化しておき,光照射により局所で活性化する技術)により,単一の樹状突起スパインでCaMKIIの活性化を観察すると,CaMKIIの活性化はごく短期間であり,刺激後1分以内に基線レベルまで戻った(図4)32–34).T286A変異体を用いると,活性化の持続時間はさらに短くなり,T286のリン酸化が寄与することは確認されたが,それでもLismanのモデル17)から予想されるような,持続的な活性化は観察されなかった.さらに,燕麦の向光性に関与する光感受性タンパク質のLOV2ドメインを用いて作製した,光活性型CaMKII阻害ペプチドphoto-activatable AIP(PA-AIP2)を用い,LTP誘導時と誘導後1分後にそれぞれAIPを活性化させたところ,誘導時に活性化したときにはLTPは阻害されたが,1分遅らせたのみで抑制されなくなった35, 36).一方で,CaMKIIの細胞内濃度は非常に高く,単量体換算で20~80 µMとされており32),光活性化されたPA-AIP2がはたして十分にCaMKIIを抑制することができるかは不明である.
CaMKIIは多機能性リン酸化酵素であり,数百に及ぶ基質が知られている(Phosphosite Plusデーターベース:https://www.phosphosite.org/).それがLTPに必要なCaMKIIの基質を同定するのを難しくしている.
よく知られている基質にAMPA受容体がある.LTP誘導により,AMPA受容体のカルボキシ末端に存在するセリン(S)831がCaMKIIによりリン酸化され37–42),その結果受容体の活性が上がるという図式が想定されてきた43).実際に,LTPに伴いチャネルのコンダクタンスの上昇が観察されている44).この部位のノックイン動物も作製され,LTPの減少と記憶・学習障害が認められた45).
ところが,その後,グルタミン酸受容体の構造上46),リン酸化部位S831は受容体のチャネルとは離れた部位に存在し,この図式に疑問が持たれた.我々がPhos-tag SDS-PAGE47)を用い,リン酸化される受容体の割合を調べたところ,リン酸化されているAMPA受容体の量は1%以下と非常に少なく,これまでの図式は成り立たないことがわかった48).ある程度の割合の受容体がリン酸化されている前提で話が考えられてきたが,皮肉なことに,よく使われる抗リン酸化AMPA受容体抗体の感度が十分高かったため,実際には非常に低い割合のリン酸化を検出していたわけである.今後はリン酸化されるタンパク質の絶対量を議論する必要がある.
一方で,AMPA受容体の補助サブユニットであるStargazin[およびそれを含む一群のタンパク質transmembrane AMPA receptor regulatory proteins(TARP)]はCaMKIIによって,高い割合でリン酸化されることが知られている48–50).このリン酸化の意義は議論があり,リン脂質との相互作用を促すことで,AMPA受容体の細胞表面での分布と安定性を変化させる一方49, 50),シナプス後部足場タンパク質であるPSD-95との相互作用を減弱させるという報告もある51).
SynGAPは低分子Gタンパク質であるRasの不活性化因子である.シナプス後膜肥厚(post-synaptic densities:PSD)ではCaMKIIに次いで発現量の多いタンパク質である52, 53).SynGAPはCaMKIIの活性化によりシナプスから離散し,その結果,Rasが活性化される54).RasはAMPA受容体をシナプスに移行させる活性があり,それによりLTPが起こるという考えが提唱されている55).実際,FRETセンサーを用いた研究から,RasがLTP誘導後シナプスで20分程度は活性を保ち続けることがわかっている56).
他にもCaMKIIの基質は多数あり,どれがLTPに重要であるかという決着はまだついていない.
4. CaMKIIとGluN2Bの結合によるCaMKIIの活性化とシナプス移行
かねてからCaMKIIには可溶型と顆粒型(膜画分に結合している型)があることが知られてきた57).実験動物の断頭後の直後よりも1時間後に調製した膜画分,特にPSDに結合するCaMKIIが増加することも見いだされてきた58).当初は,虚血による非生理学現象と思われてきた.ところが同様な現象が,精製したCaMKIIとPSDや脳スライス標品でも再現され,しかもこれがCaMKIIの自己リン酸化によって制御されていることがわかった59).GFPを融合したCaMKIIでの生細胞イメージング実験で,LTPに伴いCaMKIIが自己リン酸化依存的にシナプスへ移行することが見事に実証された60–62).
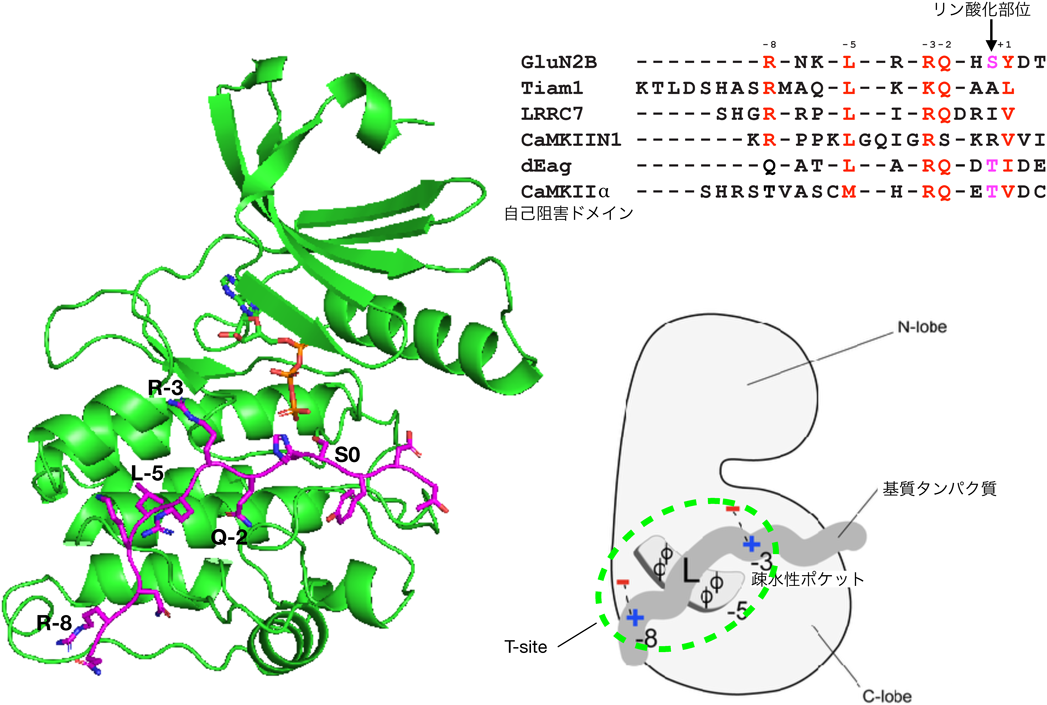
CaMKIIが神経活動依存的にシナプスに集積するのであれば,PSDタンパク質のどれかに結合していると考えられる.Colbranらはゲルオーバーレイアッセイにより,190 kDa付近にCaMKIIの結合タンパク質があることを見いだし,それがNMDA受容体GluN2B(NR2B)サブユニットであることを示した(図5)63).この結合により,CaMKIIがNMDA受容体の直下に置かれることになり,カルシウム流入がより効率よくCaMKIIを活性化する機構であると想定された.興味深いことにCaMKIIとの結合はGluN2Bサブユニット特異的で,GluN2AやGluN1など他のサブユニットとは結合しない63).GluN2BのCaMKII結合部位をノックインにより変異させると,CaMKIIの活動依存性のシナプス移行が阻害されると同時に,シナプス可塑性の異常,ならびに記憶学習の障害も観察された64).
BayerらはGluN2BとCaMKIIの結合をより詳細に解析し,GluN2BのCaMKIIとの相互作用部位はCaMKIIの自己阻害ドメインと相同性があり(図6),同じ部位(T286周囲の配列であることからT-siteと呼ばれる)に結合すると考えられた65).我々は最近GluN2Bに結合した状態のCaMKIIの結晶化に成功している(図6)66).
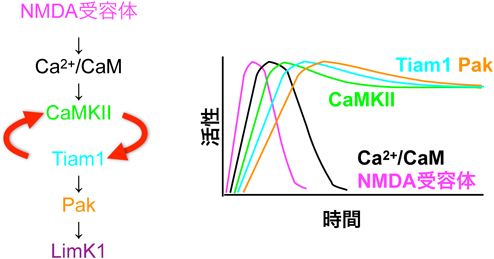
興味深いことに,この結合により,CaMKIIはCa2+/カルモジュリン非存在下でも活性化状態を保った.すなわち,T-siteにGluN2Bが結合することで,自己阻害が機能しなくなり,一部のCaMKIIがシナプス直下で活性を保つと考えられる.これは,一見前述したCaMKIIのFRETセンサーを用いてCaMKIIの活性化が一時的であるというLeeらの実験の結果(図4)32)と整合性がないが,CaMKIIは一つのPSDに5600分子(単量体換算),一方NMDA受容体は20分子と53),量が圧倒的に違うため,NMDA受容体に結合することができず,結合して活性型になるのはごく一部と予想される.また,Leeらは,基線レベルを原点として刺激前との差をみているが,刺激前のCaMKII活性の有無には言及していない.そのため,刺激前のシナプスでCaMKIIの活性があることも十分考えられる.この点は,より詳細な研究が必要となる.
5. reciprocally-activating kinase-effector complex(RAKEC)
さらに實吉らは低分子Gタンパク質Racの活性化因子(グアニンヌクレオチド交換因子GEF)であるTiam1もGluN2Bと同様,Ca2+/カルモジュリン依存性にCaMKIIのT-siteに結合し(図6),CaMKIIの活性を持続させることを見いだした(図7)34).Tiam1はCaMKIIによりリン酸化され活性化するため,Tiam1とCaMKIIの複合体は,相互に活性化させることで,その活性を持続させるメカニズムではないかと考え,これをreciprocally-activating kinase-effector complex(RAKEC)と名づけた.FRETを用い,CaMKIIとTiam1の結合をモニターしたところ,LTP誘導から少なくとも30分にわたって継続した34).このメカニズムにより,一過性のカルシウムが持続的な生化学的シグナルに変換されると考えられ,細胞内シグナルの一種の特異点であるといえる.これと一致して,下流のRacの活性化も持続した34, 67).実際にTiam1のCaMKII結合部位の変異のノックインマウスは学習障害を示した68).
それでは,RAKECはさらに一般化できるであろうか.GluN2BとTiam1以外にもCaMKIIのT-siteに結合するタンパク質はEther à-go-go(eag)potassium channel(Drosophila),GJD2/コネキシン36, LRRC7/densin-180, endogenous CaMKII inhibitor peptide(CaMKIIN),L型Ca2+チャネル,低分子Gタンパク質であるRem2などいくつかが同定されている(図6)69–74).CaMKIIの圧倒的な発現量を鑑みれば,さらに他にもCaMKIIのT-siteと結合するタンパク質があると予想される.
6. CaMKIIによって起こされる液–液相分離(LLPS)
CaMKIIはキナーゼとしては非典型的な性質を持つ.まず,量が非常に多く,たとえば海馬では全タンパク質の2%,PSDの10~20%を占める53, 75, 76).この量は,アクチンやチューブリンなど,細胞骨格タンパク質に匹敵する.キナーゼであれば,酵素反応により1分子のキナーゼが多数の基質をリン酸化することができるので,こんなに多量にある必要はない.また特徴的な回転対称型の12量体構造をしている(図3)77–79).キナーゼがこのような形態を持つ必要はなく,何らかの構造的な役割があるのではないかと考えられた4).
最近,細胞内の非膜オルガネラの形成に液–液相分離(liquid–liquid phase separation:LLPS)の関与が注目されている.LLPSとはソフトマター物理学でかねてからの研究対象であり,特に生物学的なLLPSではタンパク質やRNAなどの高分子化合物が,囲む膜を必要とせず,自発的に濃集相を形成する現象である.最近LLPSが,核小体,タンパク質–RNA複合体,ストレス顆粒など数多くの生命科学的現象に関わると考えられるようになった80, 81).シナプスへのタンパク質の集積にも関与すると考えられ,シナプス顆粒タンパク質のシナプシン,PSD足場タンパク質であるSynGAP1, PSD-95, Homer, Shank, GKAPなどもLLPSを起こすことが報告された82–85).
LLPSを起こしやすいタンパク質にはいくつかの性質が知られている80, 81).一つは多価の相互作用であり,多量体形成や,複数の結合ドメインを持つ場合が相当する.もう一つは,特定の立体構造を持たない天然変性領域を持つことである.天然変性領域にはセリン,グルタミン,グルタミン酸,アルギニン,リシンなどの電荷を持つアミノ酸が多い一方,バリン,ロイシン,イソロイシンなどの疎水性アミノ酸は少ない.そのため,これらの残基による静電気的相互作用がLLPSに寄与する.
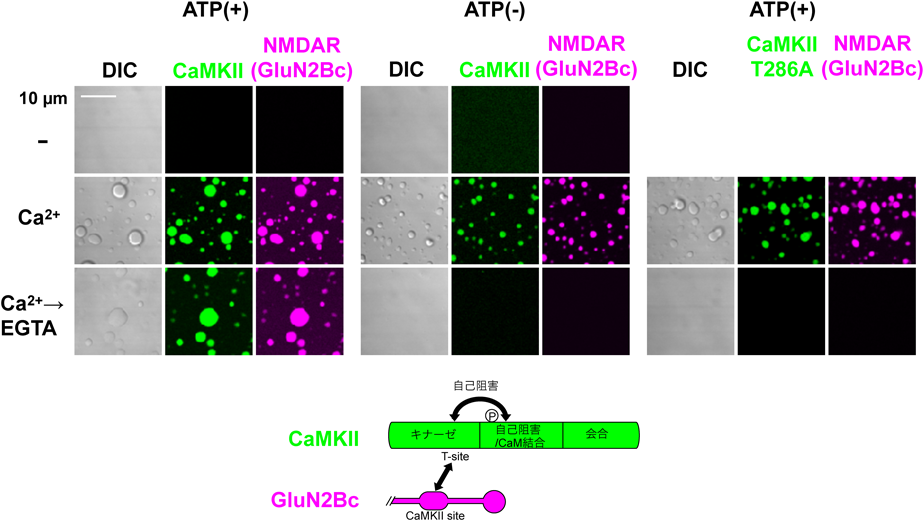
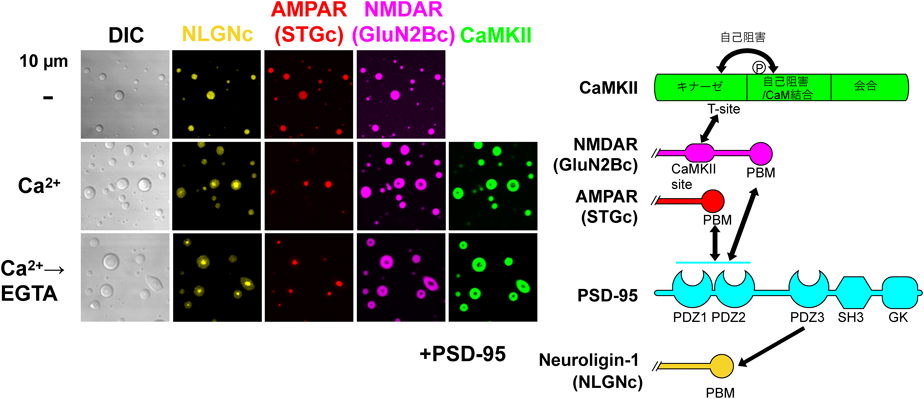
我々は,CaMKIIの多量に存在する点,12量体タンパク質であるという点,さらにGluN2BやTiam1といった一部の基質と安定した複合体を作る点がLLPSを起こすのに最適な性質を持っているのではないかと考えた.CaMKIIの結合するGluN2Bの細胞内カルボキシ末端は,ほぼ全領域が天然変性領域である点もこれと矛盾しない.そのため,細川,劉らはCaMKIIとGluN2Bのカルボキシ末端(実際の受容体のオリゴマー構造を模倣するため,二量体の蛍光タンパク質と融合した)を精製した上で,混合し,蛍光顕微鏡下で観察した.Ca2+の非存在下ではいずれのタンパク質もLLPSを起こさなかった(図8)86).しかし,Ca2+/カルモジュリンで刺激すると,タンパク質のボール状の液滴が観察され,LLPSが起こったことが示唆された.しかも,いったん起こった相分離はEGTAでCa2+をキレートしても継続した.CaMKIIがLLPSを起こすのには,キナーゼ活性は必要なかったが,それがEGTA添加後にも継続するためにはT286の自己リン酸化が必要であった.T286A変異体や触媒部位に変異を入れた場合は,Ca2+を添加するとLLPSが起こるが,EGTAを加えると消失した.
神経細胞に発現したGFP-CaMKIIを,シナプスで蛍光退色させると数分で回復することから,シナプスのCaMKIIはシナプスに濃縮されてはいても,動的な状態にあることが示唆されていた62, 87).このことは実際にPSDのCaMKIIがLLPS状態であることを示唆している.これらの結果から,CaMKIIのシナプスでの意義は,Ca2+依存性にLLPSを起こすことにあるのではないかと考えられた.
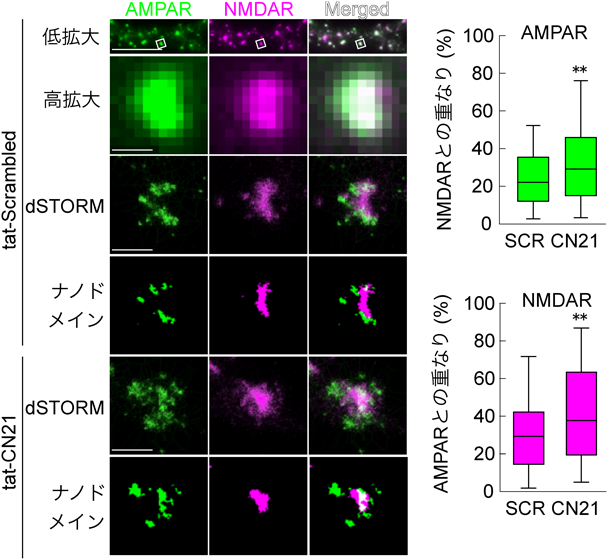
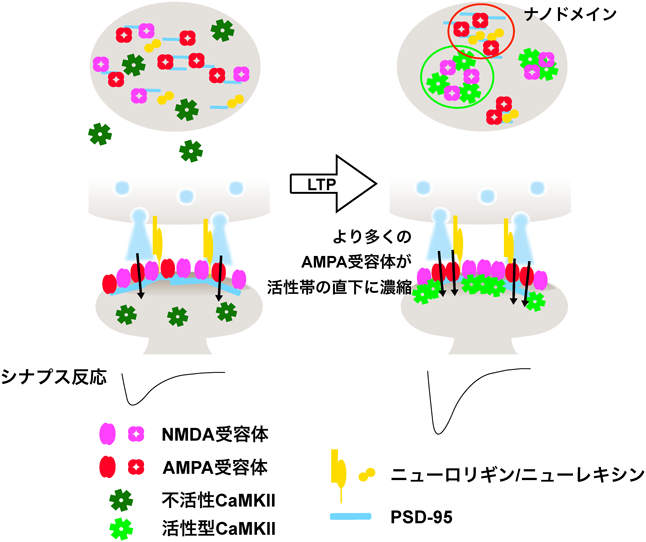
さらに我々は代表的なPSD足場タンパク質である,PSD-95, AMPA受容体補助サブユニットStargazinの細胞内カルボキシ末端,シナプス接着因子ニューロリギンの細胞内カルボキシ末端をCaMKII, GluN2B細胞内カルボキシ末端,カルモジュリンとともにLLPSを形成するかを検討した.Ca2+の非存在下では,CaMKII以外のタンパク質がLLPSを起こした(図9)86).これは足場タンパク質としてのPSD-95の役割によると考えられた.ここにCa2+を添加するとCaMKIIが濃縮相に加わった.さらに興味深いことに,濃縮相がさらに二つに分離した(相内相形成).CaMKIIとGluN2Bがともに外側の相を形成し,Stargazin, PSD-95,ニューロリギンが内側の相を形成した.実際に超高解像顕微鏡による初代培養神経細胞のシナプスにおけるPSD内のタンパク質分布を観察すると,AMPA受容体とNMDA受容体は互いに強く分離して存在する.さらに,CaMKIIとGluN2Bとの相互作用をCN21処理により抑制すると,AMPA受容体とNMDA受容体の分離が減少した(図10)86).このことは,CaMKIIがPSD内部のタンパク質の分布を制御していることを示唆している.ニューロリギンは,シナプス前部のニューレキシンと相互作用する88).ニューレキシンはシナプス顆粒の放出部位である,活性帯の構成タンパク質とも相互作用するので,このメカニズムにより,AMPA受容体がシナプス顆粒の放出部位直下に濃縮される可能性がある.AMPA受容体は,シナプスのグルタミン酸に対して飽和していない89–91).また,シナプス前部から放出されたグルタミン酸はシナプス間隙でごく局所にしか広がらない.そのため,活性化されたCaMKIIによってAMPA受容体がシナプス顆粒放出部位直下に濃縮されることにより,AMPA受容体活性が増加することが,シナプス可塑性の一つのメカニズムである可能性がある(図11)92).
LLPSには外部のタンパク質を選別し取り込んだり排斥したりする性質があり,取り込まれるタンパク質はクライアントと呼ばれる.すなわち,LTPの誘導に伴い,シナプスのGluN2BとCaMKIIが相互作用し,シナプスでLLPSを起こすと,それに対してクライアントタンパク質が取り込まれると考えられる.これにより,LTPに伴うシナプスの再構成が起こると考えられる62).実際にどのようなクライアントがあるかどうかが今後の重要な課題になっていくであろう.
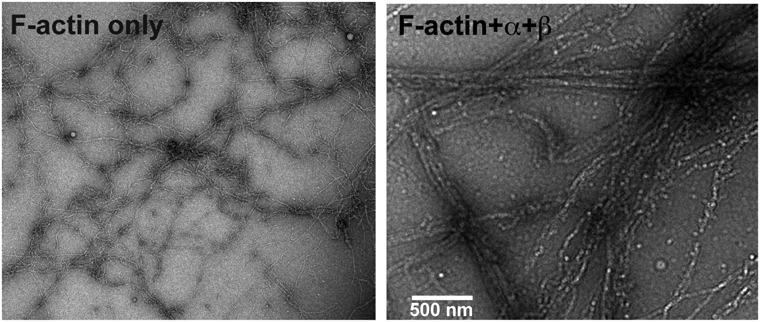
CaMKIIサブユニットのうち,β, γ, δサブユニットには調節ドメインと会合ドメインの間に特有の配列がある.この領域は天然変性領域であり,線維状アクチン(F-アクチン)と結合することが知られている(図3A,緑)93).前脳ではαとβサブユニットが平均約3:1の割合で共発現している.CaMKIIは12量体であるため,一つのホロ酵素(オリゴマー)に平均して3分子程度のβサブユニットを含んでいると考えられる8).そのため,一つのホロ酵素が複数のF-アクチンと同時に相互作用し,これにより単に結合するだけではなく,F-アクチンを束化するのではないかと考えられた.我々はこれを実証するため,電子顕微鏡観察(図12)と生化学的解析を行い,CaMKIIβサブユニットがF-アクチンを束化することを確認した94, 95).さらにサブユニット特異的なshRNAを神経細胞で用いたところ,βサブユニットに対するshRNAを用いたときにのみスパイン形態が縮小した.一方,αサブユニットに対するshRNAでは変化がなかった94).このスパインの縮小は,βサブユニットの野生型だけではなく,キナーゼ活性を失わせた変異体でもレスキューできたことから,CaMKIIの酵素活性が必要なのではなく,F-アクチン束化活性が重要であることが示唆された94).
アクチン結合部位には,多数のセリン,トレオニンが存在し,そのほとんどが自己リン酸化され,それによりF-アクチンとの結合が減弱する(図3A)96).FRETを用い,CaMKIIβサブユニットとF-アクチンとの結合をモニターしたところ,LTP誘導直後に両者は乖離し,1分程度で再結合することがわかった97).この間に,コフィリンやArp2/3などのアクチン調節因子がアクチンに作用し,スパインの形態を変化させる.この過程をセリン,トレオニンをアラニンに変異させて阻害すると,LTPも阻害された97).
アクチン束化因子としてのCaMKIIの役割にLLPSがどのように関与しているかはまだ不明であり,今後の研究の進展が待たれる.
近年のエクソームシーケンシングの発展により,ヒトにおいてCaMKIIの遺伝変異が見いだされた.秋田らは976人の知的能力障害(知的発達症)の全エクソーム解析を行い,3例のCaMKIIαサブユニット,2例のCaMKIIβサブユニットのde novo変異を見いだしている98).またKüryらはヨーロッパならびにアメリカで見つかったそれぞれ12例,7例のde novo変異99),さらに1例のCaMKIIγサブユニットの変異100)を,ChiaらもヨルダンのCaMKIIαサブユニット変異1家系2例を報告している101).いずれも患者は臨床的には重篤な知的能力障害,症例によっては痙攣や,画像上,脳萎縮を呈する例もある.またIossifovらは自閉症を示す患者のエクソーム解析からCaMKIIαサブユニット変異を1例見いだしている102).
これらは主にコーディング領域の変異であるが,イントロンのスプライス領域と考えられる変異も見つかっている.コーディング領域の変異は,キナーゼドメインにあるもの,自己阻害ドメインにあるもの,会合ドメインにあるものなど多岐にわたる.興味深いことに,CaMKIIαサブユニットのPro212, Pro235の変異は秋田ら,Küryらのいずれにも独立して見いだされており,CaMKII機能へ重要であることが示唆される.また自閉症患者でみられたCaMKIIαのE183V変異体は,E183V変異もKüryらによっても検出されている.この変異はノックイン動物も作られ,詳細に解析されている103).
我々にとって興味深いのは,F98S変異である99).F98は,BayerらによりT-siteを構成しGluN2Bとの結合に重要なアミノ酸残基として同定されており,CaMKIIのLLPSを阻害することが予想される.したがって,ヒトでの変異により重篤な症状が出たということは,CaMKIIのLLPS形成能がその機能に重要であること示している.
謝辞Acknowledgments
本稿で紹介した研究に関わった多くの共同研究者の方々に感謝いたします.また,溝口萌さん,白川日菜実さん,杉山優衣さんにはコメントをいただきましたことを感謝いたします.理研理事長ファンド,京都大学SPIRITS 2019, 科研費18H05434, 20K21462, 上原記念財団,内藤記念科学振興財団,武田科学振興財団,日本応用酵素協会,Human Frontier Science Foundationの援助を得て行われました.この場をお借りしてお礼申し上げます.
引用文献References
1) ヘッブ,D.O.(2011)行動の機構 脳メカニズムから心理学へ.岩波書店,東京.
2) Bliss, T.V. & Lømo, T. (1973) Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol., 232, 331–356.
3) Bliss, T.V. & Collingridge, G.L. (1993) A synaptic model of memory: Long-term potentiation in the hippocampus. Nature, 361, 31–39.
4) Kim, K., Saneyoshi, T., Hosokawa, T., Okamoto, K., & Hayashi, Y. (2016) Interplay of enzymatic and structural functions of CaMKII in long-term potentiation. J. Neurochem., 139, 959–972.
5) Yamauchi, T. & Fujisawa, H. (1983) Purification and characterization of the brain calmodulin-dependent protein kinase (kinase II), which is involved in the activation of tryptophan 5-monooxygenase. Eur. J. Biochem., 132, 15–21.
6) 藤澤 仁(1992)多機能性カルモジュリン依存性タンパク質リン酸化酵素.生化学,64, 14–25.
7) 山内 卓(2007)カムキナーゼIIから記憶・学習の分子的基盤へ.Yakugaku Zasshi, 127, 1173–1197.
8) Bennett, M.K., Erondu, N.E., & Kennedy, M.B. (1983) Purification and characterization of a calmodulin-dependent protein-kinase that is highly concentrated in brain. J. Biol. Chem., 258, 2735–2744.
9) Kuret, J. & Schulman, H. (1984) Purification and characterization of a Ca2+/calmodulin-dependent protein kinase from rat brain. Biochemistry, 23, 5495–5504.
10) Schworer, C.M., McClure, R.W., & Soderling, T.R. (1985) Calmodulin-dependent protein kinases purified from rat brain and rabbit liver. Arch. Biochem. Biophys., 242, 137–145.
11) Kelly, P.T., McGuinness, T.L., & Greengard, P. (1984) Evidence that the major postsynaptic density protein is a component of a Ca2+/calmodulin-dependent protein kinase. Proc. Natl. Acad. Sci. USA, 81, 945–949.
12) Tobimatsu, T. & Fujisawa, H. (1989) Tissue-specific expression of four types of rat calmodulin-dependent protein kinase II mRNAs. J. Biol. Chem., 264, 17907–17912.
13) Brocke, L., Srinivasan, M., & Schulman, H. (1995) Developmental and regional expression of multifunctional Ca2+/calmodulin-dependent protein kinase isoforms in rat brain. J. Neurosci., 15, 6797–6808.
14) Cook, S.G., Bourke, A.M., O’Leary, H., Zaegel, V., Lasda, E., Mize-Berge, J., Quillinan, N., Tucker, C.L., Coultrap, S.J., Herson, P.S., et al. (2018) Analysis of the CaMKIIalpha and beta splice-variant distribution among brain regions reveals isoform-specific differences in holoenzyme formation. Sci. Rep., 8, 5448.
15) Schworer, C.M., Colbran, R.J., & Soderling, T.R. (1986) Reversible generation of a Ca2+-independent form of Ca2+(calmodulin)-dependent protein kinase II by an autophosphorylation mechanism. J. Biol. Chem., 261, 8581–8584.
16) Nishiyama, J. & Yasuda, R. (2015) Biochemical computation for spine structural plasticity. Neuron, 87, 63–75.
17) Lisman, J.E. & Goldring, M.A. (1988) Feasibility of long-term storage of graded information by the Ca2+/calmodulin-dependent protein kinase molecules of the postsynaptic density. Proc. Natl. Acad. Sci. USA, 85, 5320–5324.
18) Malinow, R., Schulman, H., & Tsien, R.W. (1989) Inhibition of postsynaptic pkc or camkii blocks induction but not expression of ltp. Science, 245, 862–866.
19) Malenka, R.C., Kauer, J.A., Perkel, D.J., Mauk, M.D., Kelly, P.T., Nicoll, R.A., & Waxham, M.N. (1989) An essential role for postsynaptic calmodulin and protein kinase activity in long-term potentiation. Nature, 340, 554–557.
20) Pettit, D.L., Perlman, S., & Malinow, R. (1994) Potentiated transmission and prevention of further ltp by increased camkii activity in postsynaptic hippocampal slice neurons. Science, 266, 1881–1885.
21) Lledo, P.M., Hjelmstad, G.O., Mukherji, S., Soderling, T.R., Malenka, R.C., & Nicoll, R.A. (1995) Calcium/calmodulin-dependent kinase II and long-term potentiation enhance synaptic transmission by the same mechanism. Proc. Natl. Acad. Sci. USA, 92, 11175–11179.
22) Shirke, A.M. & Malinow, R. (1997) Mechanisms of potentiation by calcium-calmodulin kinase II of postsynaptic sensitivity in rat hippocampal CA1 neurons. J. Neurophysiol., 78, 2682–2692.
23) Hayashi, Y., Shi, S.H., Esteban, J.A., Piccini, A., Poncer, J.C., & Malinow, R. (2000) Driving AMPA receptors into synapses by LTP and CaMKII: Requirement for GluR1 and PDZ domain interaction. Science, 287, 2262–2267.
24) Silva, A.J., Stevens, C.F., Tonegawa, S., & Wang, Y. (1992) Deficient hippocampal long-term potentiation in alpha-calcium-calmodulin kinase II mutant mice. Science, 257, 201–206.
25) Silva, A.J., Wang, Y., Paylor, R., Wehner, J.M., Stevens, C.F., & Tonegawa, S. (1992) Alpha calcium/calmodulin kinase II mutant mice: Deficient long-term potentiation and impaired spatial learning. Cold Spring Harb. Symp. Quant. Biol., 57, 527–539.
26) Giese, K.P., Fedorov, N.B., Filipkowski, R.K., & Silva, A.J. (1998) Autophosphorylation at Thr286 of the alpha calcium-calmodulin kinase II in LTP and learning. Science, 279, 870–873.
27) Ouyang, Y., Kantor, D., Harris, K.M., Schuman, E.M., & Kennedy, M.B. (1997) Visualization of the distribution of autophosphorylated calcium/calmodulin-dependent protein kinase II after tetanic stimulation in the CA1 area of the hippocampus. J. Neurosci., 17, 5416–5427.
28) Fukunaga, K., Stoppini, L., Miyamoto, E., & Muller, D. (1993) Long-term potentiation is associated with an increased activity of Ca2+ calmodulin-dependent protein kinase-ii. J. Biol. Chem., 268, 7863–7867.
29) Fukunaga, K., Muller, D., & Miyamoto, E. (1995) Increased phosphorylation of Ca2+/calmodulin-dependent protein kinase II and its endogenous substrates in the induction of long-term potentiation. J. Biol. Chem., 270, 6119–6124.
30) Takao, K., Okamoto, K., Nakagawa, T., Neve, R.L., Nagai, T., Miyawaki, A., Hashikawa, T., Kobayashi, S., & Hayashi, Y. (2005) Visualization of synaptic Ca2+/calmodulin-dependent protein kinase II activity in living neurons. J. Neurosci., 25, 3107–3112.
31) Kwok, S., Lee, C., Sanchez, S.A., Hazlett, T.L., Gratton, E., & Hayashi, Y. (2008) Genetically encoded probe for fluorescence lifetime imaging of CaMKII activity. Biochem. Biophys. Res. Commun., 369, 519–525.
32) Lee, S.J., Escobedo-Lozoya, Y., Szatmari, E.M., & Yasuda, R. (2009) Activation of CaMKII in single dendritic spines during long-term potentiation. Nature, 458, 299–304.
33) Chang, J.Y., Parra-Bueno, P., Laviv, T., Szatmari, E.M., Lee, S.R., & Yasuda, R. (2017) CaMKII autophosphorylation is necessary for optimal integration of Ca2+ signals during LTP induction, but not maintenance. Neuron, 94, 800–808.e804.
34) Saneyoshi, T., Matsuno, H., Suzuki, A., Murakoshi, H., Hedrick, N.G., Agnello, E., O’Connell, R., Stratton, M.M., Yasuda, R., & Hayashi, Y. (2019) Reciprocal activation within a kinase-effector complex underlying persistence of structural LTP. Neuron, 102, 1199–1210.e1196.
35) Ishida, A., Kameshita, I., Okuno, S., Kitani, T., & Fujisawa, H. (1995) A novel highly specific and potent inhibitor of calmodulin-dependent protein kinase II. Biochem. Biophys. Res. Commun., 212, 806–812.
36) Murakoshi, H., Shin, M.E., Parra-Bueno, P., Szatmari, E.M., Shibata, A.C., & Yasuda, R. (2017) Kinetics of endogenous CaMKII required for synaptic plasticity revealed by optogenetic kinase inhibitor. Neuron, 94, 37–47.e35.
37) McGlade-McCulloh, E., Yamamoto, H., Tan, S.E., Brickey, D.A., & Soderling, T.R. (1993) Phosphorylation and regulation of glutamate receptors by calcium/calmodulin-dependent protein kinase II. Nature, 362, 640–642.
38) Barria, A., Muller, D., Derkach, V., Griffith, L.C., & Soderling, T.R. (1997) Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science, 276, 2042–2045.
39) Barria, A., Derkach, V., & Soderling, T. (1997) Identification of the Ca2+/calmodulin-dependent protein kinase II regulatory phosphorylation site in the α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate-type glutamate receptor. J. Biol. Chem., 272, 32727–32730.
40) Mammen, A.L., Kameyama, K., Roche, K.W., & Huganir, R.L. (1997) Phosphorylation of the alpha-amino-3-hydroxy-5-methylisoxazole4-propionic acid receptor GluR1 subunit by calcium/calmodulin-dependent kinase II. J. Biol. Chem., 272, 32528–32533.
41) Roche, K.W., O’Brien, R.J., Mammen, A.L., Bernhardt, J., & Huganir, R.L. (1996) Characterization of multiple phosphorylation sites on the AMPA receptor GluR1 subunit. Neuron, 16, 1179–1188.
42) Lee, H.K., Barbarosie, M., Kameyama, K., Bear, M.F., & Huganir, R.L. (2000) Regulation of distinct AMPA receptor phosphorylation sites during bidirectional synaptic plasticity. Nature, 405, 955–959.
43) Derkach, V., Barria, A., & Soderling, T.R. (1999) Ca2+/calmodulin-kinase II enhances channel conductance of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate type glutamate receptors. Proc. Natl. Acad. Sci. USA, 96, 3269–3274.
44) Benke, T.A., Luthi, A., Isaac, J.T., & Collingridge, G.L. (1998) Modulation of AMPA receptor unitary conductance by synaptic activity. Nature, 393, 793–797.
45) Lee, H.K., Takamiya, K., Han, J.S., Man, H., Kim, C.H., Rumbaugh, G., Yu, S., Ding, L., He, C., Petralia, R.S., et al. (2003) Phosphorylation of the AMPA receptor GluR1 subunit is required for synaptic plasticity and retention of spatial memory. Cell, 112, 631–643.
46) Armstrong, N., Sun, Y., Chen, G.Q., & Gouaux, E. (1998) Structure of a glutamate-receptor ligand-binding core in complex with kainate. Nature, 395, 913–917.
47) Kinoshita, E., Kinoshita-Kikuta, E., Matsubara, M., Yamada, S., Nakamura, H., Shiro, Y., Aoki, Y., Okita, K., & Koike, T. (2008) Separation of phosphoprotein isotypes having the same number of phosphate groups using phosphate-affinity SDS-PAGE. Proteomics, 8, 2994–3003.
48) Hosokawa, T., Mitsushima, D., Kaneko, R., & Hayashi, Y. (2015) Stoichiometry and phosphoisotypes of hippocampal AMPA type glutamate receptor phosphorylation. Neuron, 85, 60–67.
49) Sumioka, A., Yan, D., & Tomita, S. (2010) TARP phosphorylation regulates synaptic AMPA receptors through lipid bilayers. Neuron, 66, 755–767.
50) Opazo, P., Labrecque, S., Tigaret, C.M., Frouin, A., Wiseman, P.W., De Koninck, P., & Choquet, D. (2010) CaMKII triggers the diffusional trapping of surface AMPARs through phosphorylation of stargazin. Neuron, 67, 239–252.
51) Zeng, M., Diaz-Alonso, J., Ye, F., Chen, X., Xu, J., Ji, Z., Nicoll, R.A., & Zhang, M. (2019) Phase separation-mediated TARP/MAGUK complex condensation and AMPA receptor synaptic transmission. Neuron, 104, 529–543.e526.
52) Cheng, D., Hoogenraad, C.C., Rush, J., Ramm, E., Schlager, M.A., Duong, D.M., Xu, P., Wijayawardana, S.R., Hanfelt, J., Nakagawa, T., et al. (2006) Relative and absolute quantification of postsynaptic density proteome isolated from rat forebrain and cerebellum. Mol. Cell. Proteomics, 5, 1158–1170.
53) Sheng, M. & Hoogenraad, C.C. (2007) The postsynaptic architecture of excitatory synapses: A more quantitative view. Annu. Rev. Biochem., 76, 823–847.
54) Araki, Y., Zeng, M., Zhang, M., & Huganir, R.L. (2015) Rapid dispersion of SynGAP from synaptic spines triggers AMPA receptor insertion and spine enlargement during LTP. Neuron, 85, 173–189.
55) Zhu, J.J., Qin, Y., Zhao, M., Van Aelst, L., & Malinow, R. (2002) Ras and Rap control AMPA receptor trafficking during synaptic plasticity. Cell, 110, 443–455.
56) Harvey, C.D., Yasuda, R., Zhong, H., & Svoboda, K. (2008) The spread of Ras activity triggered by activation of a single dendritic spine. Science, 321, 136–140.
57) Yamauchi, T., Sekihara, S., & Ohsako, S. (1990) Subcellular distribution of alpha and beta subunit proteins of Ca2+/calmodulin-dependent protein kinase II expressed in Chinese hamster ovary cells. FEBS Lett., 266, 55–58.
58) Suzuki, T., Okumura-Noji, K., Tanaka, R., & Tada, T. (1994) Rapid translocation of cytosolic Ca2+/calmodulin-dependent protein kinase II into postsynaptic density after decapitation. J. Neurochem., 63, 1529–1537.
59) Strack, S., Choi, S., Lovinger, D.M., & Colbran, R.J. (1997) Translocation of autophosphorylated calcium/calmodulin-dependent protein kinase II to the postsynaptic density. J. Biol. Chem., 272, 13467–13470.
60) Shen, K. & Meyer, T. (1999) Dynamic control of CaMKII translocation and localization in hippocampal neurons by NMDA receptor stimulation. Science, 284, 162–166.
61) Shen, K., Teruel, M.N., Connor, J.H., Shenolikar, S., & Meyer, T. (2000) Molecular memory by reversible translocation of calcium/calmodulin-dependent protein kinase II. Nat. Neurosci., 3, 881–886.
62) Bosch, M., Castro, J., Saneyoshi, T., Matsuno, H., Sur, M., & Hayashi, Y. (2014) Structural and molecular remodeling of dendritic spine substructures during long-term potentiation. Neuron, 82, 444–459.
63) Strack, S. & Colbran, R.J. (1998) Autophosphorylation-dependent targeting of calcium/calmodulin-dependent protein kinase II by the NR2B subunit of the N-methyl-D-aspartate receptor. J. Biol. Chem., 273, 20689–20692.
64) Halt, A.R., Dallapiazza, R.F., Zhou, Y., Stein, I.S., Qian, H., Juntti, S., Wojcik, S., Brose, N., Silva, A.J., & Hell, J.W. (2012) CaMKII binding to GluN2B is critical during memory consolidation. EMBO J., 31, 1203–1216.
65) Bayer, K.U., De Koninck, P., Leonard, A.S., Hell, J.W., & Schulman, H. (2001) Interaction with the NMDA receptor locks CaMKII in an active conformation. Nature, 411, 801–805.
66) Özden, C., Sloutsky, R., Santos, N., Agnello, E., Gaubitz, C., Lapinskas, E., Esposito, E.A., Kelch, B.A., Garman, S.C., Hayashi, Y., et al. (2020) CaMKII binds both substrates and effectors at the active site. bioRxiv, 10.25.354241.
67) Hedrick, N.G., Harward, S.C., Hall, C.E., Murakoshi, H., McNamara, J.O., & Yasuda, R. (2016) Rho GTPase complementation underlies BDNF-dependent homo- and heterosynaptic plasticity. Nature, 538, 104–108.
68) Kojima, H., Rosendale, M., Sugiyama, Y., Hayashi, M., Horiguchi, Y., Yoshihara, T., Ikegaya, Y., Saneyoshi, T., & Hayashi, Y. (2019) The role of CaMKII-Tiam1 complex on learning and memory. Neurobiol. Learn. Mem., 166, 107070.
69) Chang, B.H., Mukherji, S., & Soderling, T.R. (1998) Characterization of a calmodulin kinase II inhibitor protein in brain. Proc. Natl. Acad. Sci. USA, 95, 10890–10895.
70) Walikonis, R.S., Oguni, A., Khorosheva, E.M., Jeng, C.J., Asuncion, F.J., & Kennedy, M.B. (2001) Densin-180 forms a ternary complex with the α-subunit of Ca2+/calmodulin-dependent protein kinase II and α-actinin. J. Neurosci., 21, 423–433.
71) Sun, X.X., Hodge, J.J., Zhou, Y., Nguyen, M., & Griffith, L.C. (2004) The eag potassium channel binds and locally activates calcium/calmodulin-dependent protein kinase II. J. Biol. Chem., 279, 10206–10214.
72) Alev, C., Urschel, S., Sonntag, S., Zoidl, G., Fort, A.G., Hoher, T., Matsubara, M., Willecke, K., Spray, D.C., & Dermietzel, R. (2008) The neuronal connexin36 interacts with and is phosphorylated by CaMKII in a way similar to CaMKII interaction with glutamate receptors. Proc. Natl. Acad. Sci. USA, 105, 20964–20969.
73) Wang, X., Marks, C.R., Perfitt, T.L., Nakagawa, T., Lee, A., Jacobson, D.A., & Colbran, R.J. (2017) A novel mechanism for Ca2+/calmodulin-dependent protein kinase II targeting to L-type Ca2+ channels that initiates long-range signaling to the nucleus. J. Biol. Chem., 292, 17324–17336.
74) Royer, L., Herzog, J.J., Kenny, K., Tzvetkova, B., Cochrane, J.C., Marr, M.T. 2nd, & Paradis, S. (2018) The Ras-like GTPase Rem2 is a potent inhibitor of calcium/calmodulin-dependent kinase II activity. J. Biol. Chem., 293, 14798–14811.
75) Erondu, N.E. & Kennedy, M.B. (1985) Regional distribution of type II Ca2+/calmodulin-dependent protein kinase in rat brain. J. Neurosci., 5, 3270–3277.
76) Miller, S.G. & Kennedy, M.B. (1985) Distinct forebrain and cerebellar isozymes of type II Ca2+/calmodulin-dependent protein kinase associate differently with the postsynaptic density fraction. J. Biol. Chem., 260, 9039–9046.
77) Kanaseki, T., Ikeuchi, Y., Sugiura, H., & Yamauchi, T. (1991) Structural features of Ca2+/calmodulin-dependent protein kinase II revealed by electron microscopy. J. Cell Biol., 115, 1049–1060.
78) Chao, L.H., Stratton, M.M., Lee, I.H., Rosenberg, O.S., Levitz, J., Mandell, D.J., Kortemme, T., Groves, J.T., Schulman, H., & Kuriyan, J. (2011) A mechanism for tunable autoinhibition in the structure of a human Ca2+/calmodulin-dependent kinase II holoenzyme. Cell, 146, 732–745.
79) Myers, J.B., Zaegel, V., Coultrap, S.J., Miller, A.P., Bayer, K.U., & Reichow, S.L. (2017) The CaMKII holoenzyme structure in activation-competent conformations. Nat. Commun., 8, 15742.
80) Hyman, A.A., Weber, C.A., & Julicher, F. (2014) Liquid-liquid phase separation in biology. Annu. Rev. Cell Dev. Biol., 30, 39–58.
81) 白木賢太郎(2019)相分離生物学.東京化学同人.
82) Milovanovic, D., Wu, Y., Bian, X., & De Camilli, P. (2018) A liquid phase of synapsin and lipid vesicles. Science, 361, 604–607.
83) Zeng, M., Shang, Y., Araki, Y., Guo, T., Huganir, R.L., & Zhang, M. (2016) Phase transition in postsynaptic densities underlies formation of synaptic complexes and synaptic plasticity. Cell, 166, 1163–1175.e1112.
84) Zeng, M., Chen, X., Guan, D., Xu, J., Wu, H., Tong, P., & Zhang, M. (2018) Reconstituted postsynaptic density as a molecular platform for understanding synapse formation and plasticity. Cell, 174, 1172–1187.e1116.
85) Hayashi, Y., Ford, L.K., Fioriti, L., McGurk, L., & Zhang, M. (2021) Liquid–liquid phase separation in physiology and pathophysiology of nervous system. J. Neurosci., 41, 834–844.
86) Hosokawa, T., Liu, P.-W., Cai, Q., Ferreira, J.S., Levet, F., Butler, C., Sibarita, J.B., Choquet, D., Groc, L., Hosy, E., et al. CaMKII activation persistently segregates postsynaptic proteins via liquid phase separation Nat. Neurosci., in press.
87) Okamoto, K., Nagai, T., Miyawaki, A., & Hayashi, Y. (2004) Rapid and persistent modulation of actin dynamics regulates postsynaptic reorganization underlying bidirectional plasticity. Nat. Neurosci., 7, 1104–1112.
88) Futai, K., Kim, M.J., Hashikawa, T., Scheiffele, P., Sheng, M., & Hayashi, Y. (2007) Retrograde modulation of presynaptic release probability through signaling mediated by PSD-95-neuroligin. Nat. Neurosci., 10, 186–195.
89) Patneau, D.K. & Mayer, M.L. (1990) Structure–Activity-Relationships for Amino-Acid Transmitter Candidates Acting at N-Methyl-D-Aspartate and Quisqualate Receptors. J. Neurosci., 10, 2385–2399.
90) Tong, G. & Jahr, C.E. (1994) Block of glutamate transporters potentiates postsynaptic excitation. Neuron, 13, 1195–1203.
91) Liu, G., Choi, S., & Tsien, R.W. (1999) Variability of neurotransmitter concentration and nonsaturation of postsynaptic AMPA receptors at synapses in hippocampal cultures and slices. Neuron, 22, 395–409.
92) Xie, X., Liaw, J.S., Baudry, M., & Berger, T.W. (1997) Novel expression mechanism for synaptic potentiation: Alignment of presynaptic release site and postsynaptic receptor. Proc. Natl. Acad. Sci. USA, 94, 6983–6988.
93) Shen, K., Teruel, M.N., Subramanian, K., & Meyer, T. (1998) CaMKIIβ functions as an F-actin targeting module that localizes CaMKIIα/β heterooligomers to dendritic spines. Neuron, 21, 593–606.
94) Okamoto, K., Narayanan, R., Lee, S.H., Murata, K., & Hayashi, Y. (2007) The role of CaMKII as an F-actin-bundling protein crucial for maintenance of dendritic spine structure. Proc. Natl. Acad. Sci. USA, 104, 6418–6423.
95) O’Leary, H., Lasda, E., & Bayer, K.U. (2006) CaMKIIβ association with the actin-cytoskeleton is regulated by alternative splicing. Mol. Biol. Cell, 17, 4656–4665.
96) Kim, K., Lakhanpal, G., Lu, H.E., Khan, M., Suzuki, A., Kato-Hayashi, M., Narayanan, R., Luyben, T.T., Matsuda, T., Nagai, T., et al. (2015) A temporary gating of actin remodeling during synaptic plasticity consists of the interplay between the kinase and structural functions of CaMKII. Neuron, 87, 813–826.
97) Kim, K., Suzuki, A., Kojima, H., Kawamura, M., Miya, K., Abe, M., Yamada, I., Furuse, T., Wakana, S., Sakimura, K., et al. (2019) Autophosphorylation of F-actin binding domain of CaMKIIbeta is required for fear learning. Neurobiol. Learn. Mem., 157, 86–95.
98) Akita, T., Aoto, K., Kato, M., Shiina, M., Mutoh, H., Nakashima, M., Kuki, I., Okazaki, S., Magara, S., Shiihara, T., et al. (2018) De novo variants in CAMK2A and CAMK2B cause neurodevelopmental disorders. Ann. Clin. Transl. Neurol., 5, 280–296.
99) Küry, S., van Woerden, G.M., Besnard, T., Proietti Onori, M., Latypova, X., Towne, M.C., Cho, M.T., Prescott, T.E., Ploeg, M.A., Sanders, S., et al.; Undiagnosed Diseases Network; GEM HUGO; Deciphering Developmental Disorders Study. (2017) De novo mutations in protein kinase genes CAMK2A and CAMK2B cause intellectual disability. Am. J. Hum. Genet., 101, 768–788.
100) Proietti Onori, M., Koopal, B., Everman, D.B., Worthington, J.D., Jones, J.R., Ploeg, M.A., Mientjes, E., van Bon, B.W., Kleefstra, T., Schulman, H., et al. (2018) The intellectual disability-associated CAMK2G p.Arg292Pro mutation acts as a pathogenic gain-of-function. Hum. Mutat., 39, 2008–2024.
101) Chia, P.H., Zhong, F.L., Niwa, S., Bonnard, C., Utami, K.H., Zeng, R., Lee, H., Eskin, A., Nelson, S.F., Xie, W.H., et al. (2018) A homozygous loss-of-function CAMK2A mutation causes growth delay, frequent seizures and severe intellectual disability. eLife, 7, 7.
102) Iossifov, I., O’Roak, B.J., Sanders, S.J., Ronemus, M., Krumm, N., Levy, D., Stessman, H.A., Witherspoon, K.T., Vives, L., Patterson, K.E., et al. (2014) The contribution of de novo coding mutations to autism spectrum disorder. Nature, 515, 216–221.
103) Stephenson, J.R., Wang, X., Perfitt, T.L., Parrish, W.P., Shonesy, B.C., Marks, C.R., Mortlock, D.P., Nakagawa, T., Sutcliffe, J.S., & Colbran, R.J. (2017) A novel human CAMK2A mutation disrupts dendritic morphology and synaptic transmission, and causes ASD-related behaviors. J. Neurosci., 37, 2216–2233.