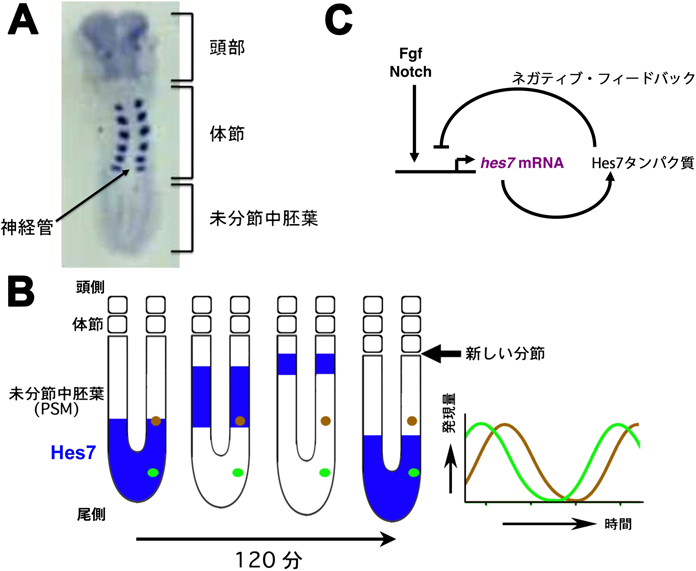
1)適切な時間遅れのネガティブ・フィードバックに依存したHes7の発現振動
体節は胎生期に一過性に形成される節状の組織で(図1A),椎骨,肋骨,骨格筋等に分化する.胎仔の尾部側にU字形をした未分節中胚葉が存在するが,その頭側部分が一定周期でくびれ切れることによって神経管の左右に1個ずつ体節が形成される(図1A, B).この周期性を決めるのは分節時計と呼ばれる生物時計で,マウスは約2時間,ヒトは約5時間周期である5–8).分節時計で中心的に働く遺伝子はHes7であり,塩基性領域・ヘリックス・ループ・ヘリックス(bHLH)構造を持つ転写抑制因子をコードする5).Hes7の発現は未分節中胚葉の尾側から始まり頭側へと移動するが,頭側の先端近くに着くと消え,その後,頭側先端部分が分節されて体節になる(図1B).マウスの場合は,このダイナミックな発現変化が約2時間周期で繰り返されるが,これは個々の未分節中胚葉細胞間でHes7が同期した発現振動をすることによる(図1B).
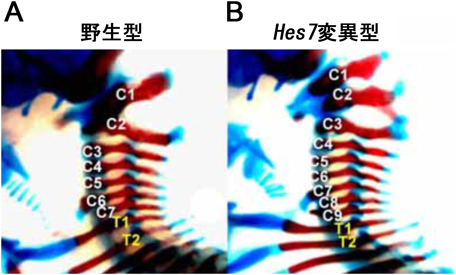
この発現振動はある程度ゆっくりとしたタイミング(適切な時間遅れ=図4A右のτ1)で起こるネガティブ・フィードバックに制御されている(図1C)9, 10).未分節中胚葉では,FgfシグナルおよびNotchシグナルによってHes7の発現が誘導されるが11),Hes7は自身のプロモーターに結合して発現を抑制する(ネガティブ・フィードバック)(図1C).Hes7 mRNAおよびタンパク質はともに非常に不安定なので,ネガティブ・フィードバックによってあらたな合成が抑制されるとすぐに分解されてなくなる.Hes7タンパク質がなくなると,ネガティブ・フィードバックが解除されて,またFgfシグナルおよびNotchシグナルによってHes7の発現が誘導される.その結果,Hes7のmRNAおよびタンパク質の発現量はともに約2時間周期で増減を繰り返す(振動).Hes7遺伝子を欠損すると,体節はすべて癒合し,体節由来の椎骨や肋骨も癒合する(図2)5).また,逆にHes7の振動を止めて一定量で持続発現させても,同様に体節の癒合が起こる10).したがって,Hes7の発現振動は分節に必須であり,Hes7は分節時計における中心的な遺伝子である.
数理モデルから,発現が振動するにはネガティブ・フィードバックはある程度ゆっくりとしたタイミングで起こる必要があり,すばやいタイミングだと定常発現になることが予測されている10, 12).これは,部屋の温度を調節するサーモスタットを考えると想像しやすい.たとえば,サーモスタットがすばやいタイミングで働けば部屋の温度は一定に保たれるが,サーモスタットがゆっくりと働くと部屋の温度は上がったり下がったりと振動する.実際にすばやいタイミングでネガティブ・フィードバックを起こす実験を行い,数理モデルの是非を確かめた.Hes7遺伝子には3個のイントロンが存在するので,転写やスプライシングにある程度の時間がかかる.Hes7遺伝子のイントロンにはエンハンサーやサイレンサーといった配列が存在しないため,イントロンがなくても発現レベルに大きな影響は与えないので,3個のイントロンをすべて除去することにした.Hes7遺伝子は転写開始からタンパク質発現まで約30分かかるが,イントロンを除去すると転写にかかる時間は短縮し,スプライシングも不要なことから,Hes7タンパク質が約20分速いタイミングで発現した.すなわち,ネガティブ・フィードバックが約20分速く起こり,そのためHes7の発現は振動せず定常になり,体節や体節由来の組織は癒合した10).したがって,ある程度ゆっくりとネガティブ・フィードバックが起こることがHes7の発現振動に必須であることが示された.
2)適切な時間遅れとイントロン数
数理モデルから,ネガティブ・フィードバックが5分程度加速すると初めのうち少し短い周期で振動するが,徐々に減衰して定常に近づくことが予測された.イントロンを2個削減すると,約5分速いタイミングでネガティブ・フィードバックが起こり,このときHes7の発現はしばらく振動するが,やがて定常になった13).興味あることに,このとき観察されたHes7の発現振動は2時間よりも短い周期で起こった.すなわち,Hes7の発現振動や分節が加速化して正常よりも多くの体節が形成され,椎骨数が増加した13).野生型では,頚部形成期にはHes7の発現は7回振動して7対の体節が作られ,その結果,頚椎骨は7個形成される(図3A).しかし,この変異マウスではHes7の発現は頚部形成期に9回振動して9対の体節が作られ,頚椎骨は9個形成された(図3B)13).これらの結果から,正しいタイミング(適切な時間遅れ)でHes7のネガティブ・フィードバックが起こることが,正常な形態形成にきわめて重要であることが明らかになった.さらにこの結果は,Hes7が分節時計における中心的なペースメーカーであることを示している.
余談になるが,上記の結果は,イントロンの新たな役割を示唆した.イントロンによってはエンハンサーやサイレンサーのような遺伝子発現制御に重要な配列を持つものもあるが,大部分のイントロンはそのような配列を持たず,ジャンク配列と呼ばれてきた.それにもかかわらず,多くの重要な遺伝子のイントロンは,その数や位置が進化上よく保存されている.イントロンの配列はあまり保存されていないのに,なぜその数や位置が保存されているのかはよくわかっていない.しかし,Hes7遺伝子から3個のイントロンを除去すると椎骨はすべて癒合し,2個のイントロンを除去すると頚椎骨は9個に増えた.特に,後者の表現型は,進化を研究する者にとっては大きな驚きであった.というのは,哺乳動物の頚椎骨は,進化上7個に保存されているからである.マウスやヒトだけでなく,首の長いキリンや首がほとんどないクジラに至るまで,哺乳動物の頚椎骨は原則7個である.しかし,ジャンク配列と思われていたイントロンを2個除去するだけで,進化上の保存が崩れたのである.Hes7遺伝子の場合,イントロンの数が0個,1個,2個以上で遺伝子発現のタイミングが5分から20分程度異なることがわかり,これが発現振動に大きな影響を与えていた.これらの結果から,イントロンの数は厳密な遺伝子発現のタイミング設定に重要な役割を担うのではないかと考えられた.Hes7以外にも,Hes1やDll1遺伝子に関してイントロンの重要性が示されてきたが(下記参照),今後,他の遺伝子についても解析が進めば,時間遅れに関するイントロンの新たな役割が明らかになっていくものと期待される.
3)適切な時間遅れの細胞間カップリングに依存した発現振動
分節過程では,未分節中胚葉細胞間でHes7は同位相で発現振動する.しかし,Notchシグナル系の変異マウスやNotchシグナル阻害剤処理において細胞間の位相関係が乱れることから,この細胞間の同位相発現振動はNotchシグナルに依存することがわかる.NotchシグナルのリガンドDelta-like1(Dll1)は発現振動するHes7によって周期的に抑制されるために,Dll1の発現も振動する.Dll1の発現が振動することによって隣接細胞のNotchシグナルが周期的に活性化され,その下流でHes7の発現も周期的に活性化される.したがって,Hes7の発現振動の情報は,Dll1の発現振動を介して隣接細胞のHes7発現振動に伝わる.実際,Dll1の発現を振動させると,隣接細胞のNotchシグナルの下流因子であるHes1の発現振動をDll1の振動に同期化できることが示された14).
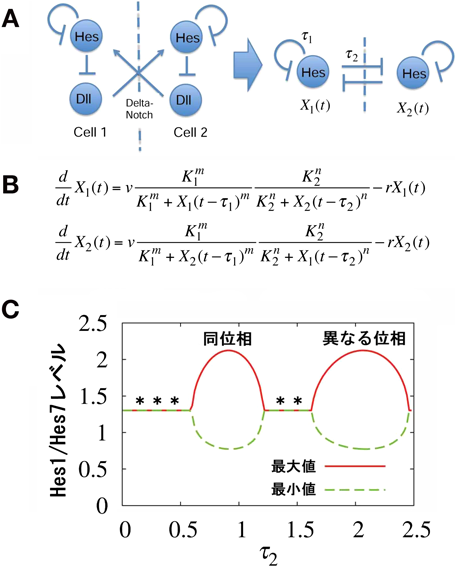
この「Hes7 ─┤ Dll1 →隣接細胞のHes7」という経路(図4A左側)は,Hes7は自分自身と隣接細胞のHes7を抑制するというダブル・ネガティブ・フィードバックに簡略化できる(図4A右側).この経路に基づく数理モデル(図4B)から,細胞間の情報伝達にかかる時間(=細胞間カップリングにおける時間遅れ=図4Aのτ2)が同位相の振動にきわめて重要であることが予測された.この時間遅れが適切な時間帯にあれば同位相の同期振動をするが,適切な時間帯から半周期程度長く,あるいは短くなると異なる位相で同期振動する(図4C)15, 16).同位相あるいは異なる位相で同期振動する時間帯の境界領域の時間遅れでは振幅が小さくなり,極端な場合は振動が停止して定常発現になる(図4C,*の領域)17).これは,oscillation deathあるいはamplitude deathと呼ばれる状態である.細胞間カップリングにおける時間遅れの重要性を検証するために,これを長くしたり短くしたりする実験を行った.
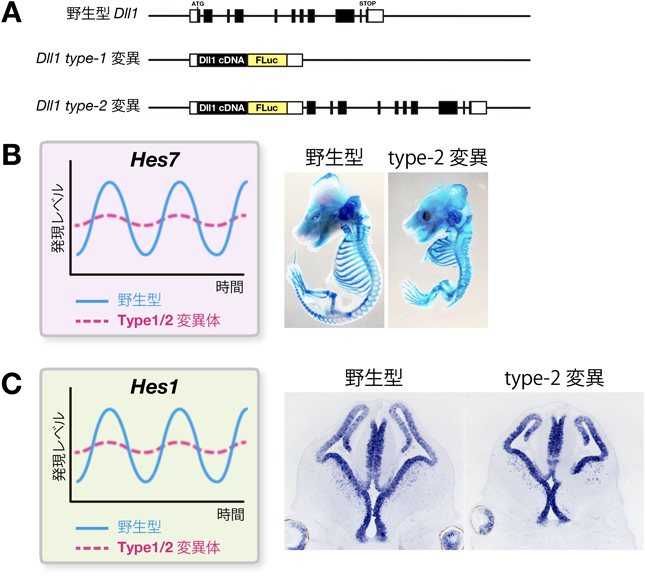
細胞間カップリングにおける時間遅れを変えるために,Dll1の発現するタイミングの加速化,あるいは遅延化を試みた.そのため,Dll1遺伝子からイントロンをすべて除去して遺伝子サイズを短くした変異(Dll1 type-1変異)および余分な配列を挿入して遺伝子サイズを長くした変異(Dll1 type-2変異)を作製した(図5A)15).前者はDll1の発現するタイミングが加速化し,後者は遅延化していた.その結果,どちらの変異でもHes7の発現振動の振幅が減少して定常発現に近づき,体節や体節由来の組織が癒合した(図5B)15).したがって,細胞間カップリングにおける時間遅れが長くなっても短くなってもoscillation deathあるいはamplitude deathと呼ばれる状態に近づくこと,すなわち正常な発現振動には適切な時間遅れを持った細胞間カップリングが必須であることが明らかになった.
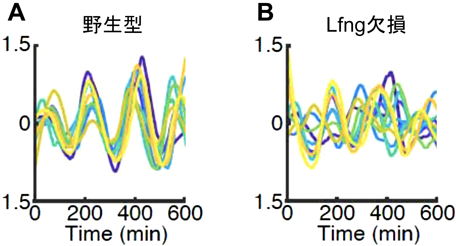
NotchシグナルのモジュレーターであるLunatic fringe(Lfng)は,Dll1やNotchに糖鎖を付加することによってそれぞれの活性を制御する.Lfngを欠損するとHes7の発現振動は影響を受けて体節が癒合するが,その詳細な分子機構はよくわかっていなかった.そこで,新たな蛍光タンパク質AchillesとHes7とを融合したタンパク質をHes7プロモーターから発現するレポーターマウスを作製したところ,Hes7タンパク質の発現をライブ・イメージングによって個々の細胞レベルで定量できるようになった16).その結果,細胞分裂で発現振動が乱れても,野生型の未分節中胚葉ではすぐに隣接細胞と同位相に同期化していた(図6A)16).しかし,Lfngを欠損すると未分節中胚葉の個々の細胞におけるHes7の発現振動の振幅が減弱するとともに,細胞間の位相関係が乱れていることがわかった(図6B)16).ところが,Lfngを欠損した未分節中胚葉細胞を単離培養すると,Hes7はほぼ正常な発現振動をしていた16).そこで,未分節中胚葉細胞間のカップリングにおける時間遅れを測定したところ,Lfng欠損細胞では正常に比べて短縮していた.この細胞間カップリングの短縮は,Lfngを欠損するとDll1タンパク質が発現してから細胞膜上に正常より速く到達するためであった16).したがって,Lfng欠損でみられる異常は,上記のDll1 type-1変異でみられる異常とよく似ていた.
以上のDll1変異やLfng欠損の解析結果から,適切な時間遅れを示す細胞間カップリングは同位相の同期振動にきわめて重要な役割を持つことが明らかになった.
1)適切な時間遅れのネガティブ・フィードバックに依存したHes1の発現振動
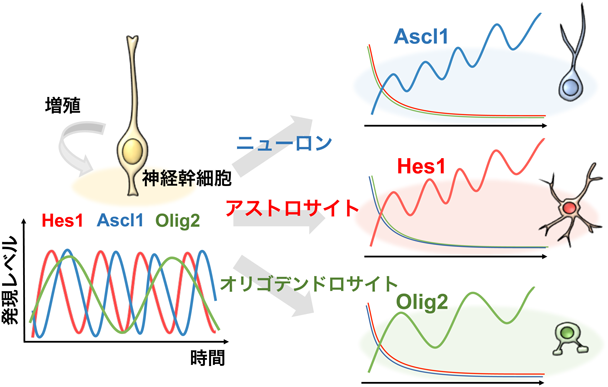
神経幹細胞は活発に増殖しつつ,経時的に分化能を変えてニューロン,オリゴデンドロサイト,アストロサイトの3種類の細胞を産生する.ニューロンへの分化は,プロニューラル因子と呼ばれるbHLH因子群によって決定される18–20).代表例としてAscl1やNeurogenin 2(Neurog2)があり,いずれもbHLH因子E47とヘリックス・ループ・ヘリックス構造を介してヘテロ二量体を形成する.このヘテロ二量体は,Eボックス(CANNTG)に結合し,ニューロン分化に必要な遺伝子群の発現を誘導する.オリゴデンドロサイトの分化は,bHLH因子Olig1やOlig2によって決定される.一方,アストロサイトの分化は,bHLH因子Hes1やHes5によって制御される.このように,これら3種類の細胞分化決定は,それぞれ特異的なbHLH因子によって制御される.しかし,Ascl1, Olig2, Hes1に関して,細胞分化決定だけでなく,未分化な神経幹細胞の増殖・維持にも働くことが示されてきた.すなわち,これらのbHLH因子は相反する活性を持つが,どのように制御されるのだろうか?
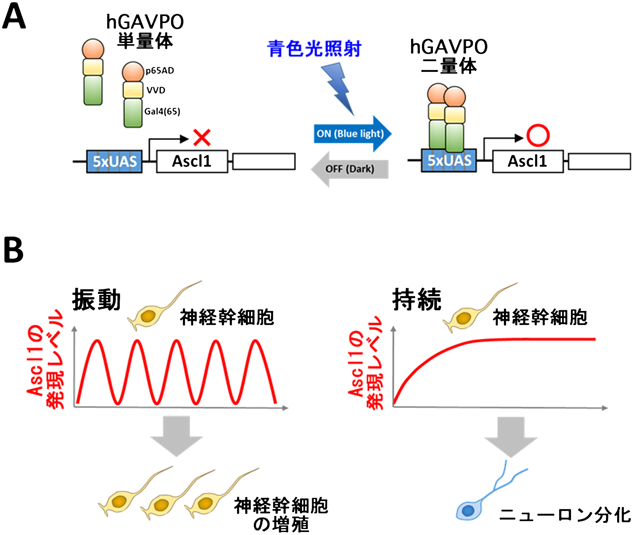
ライブ・イメージングの解析から,これらのbHLH因子群はいずれも神経幹細胞では発現が振動していた(図7)21–23).Hes1は,Hes7と同様にある程度ゆっくりとしたネガティブ・フィードバックに依存して発現振動する24, 25).Ascl1やOlig2の発現はHes1によって抑制されるので,Hes1の発現振動によって,Ascl1やOlig2の発現も振動していた.一方,ニューロン,オリゴデンドロサイト,アストロサイトの分化時には,それぞれAscl1, Olig2, Hes1が選ばれて持続発現していた(図7)22, 23).したがって,Ascl1, Olig2, Hes1はいずれも発現が振動すると神経幹細胞の増殖が活性化され,どれか一つが選ばれて持続発現すると分化決定が起こると考えられた.次に,発現動態の重要性を調べるために光遺伝学的技術を用いてAscl1の発現操作実験を行った.この光遺伝学的技術は光反応性のLOVドメインVVDを持つ因子hGAVPOを用いるもので,青色光照射でhGAVPOは活性型になり,下流遺伝子の発現を誘導する(図8A)23, 26).この技術を用いることによって,青色光照射で神経幹細胞にAscl1の発現を誘導でき,暗くするとAscl1の発現をなくせるようになった.解析の結果,Ascl1の発現が振動すると神経幹細胞の増殖が活性化されたが,Ascl1の発現が持続すると神経幹細胞は増殖を止めてニューロンに分化した(図8B)23).したがって,Ascl1は発現動態を変えることによって相反する機能を発揮することがわかった.これらの結果から,神経幹細胞の多分化能とは,Ascl1, Hes1, Olig2といった複数の分化決定因子の発現が振動することによって互いに拮抗しあった状態であるといえる.
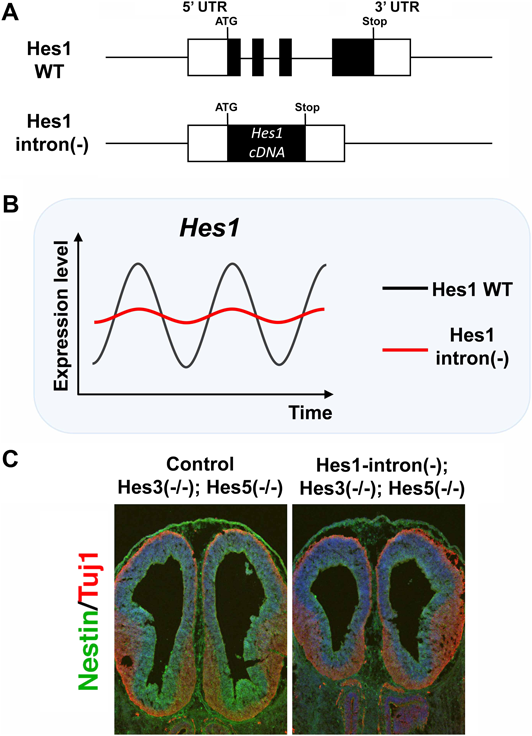
このように神経幹細胞においても遺伝子発現振動が重要であることが示された.特に,Hes1は同じファミリー分子であるHes3やHes5と一緒に神経幹細胞の増殖・維持に中心的な役割を担う.Hes1は,Hes7のように自身のプロモーターに結合して発現を抑制する(ネガティブ・フィードバック).Hes1の発現振動もゆっくりとしたタイミングのネガティブ・フィードバックに依存するかどうかを明らかにするために,Hes1の発現を加速する実験を行った25).Hes1遺伝子にも3個のイントロンが存在するので,このイントロンをすべて除去したところ,Hes1の発現が加速し,振動の振幅が減弱した(図9A, B)25).したがって,ある程度ゆっくりとネガティブ・フィードバックが起こることがHes1の発現振動に必須であることが示された.この変異マウスの脳は小さい傾向がみられたが,あまり大きな異常はみられなかった.これは,Hes1と同様の機能を持つHes3やHes5によって代償されているためと考えられた.そこで,Hes3とHes5とを欠損したマウスを使って解析した.Hes3とHes5とを欠損したマウスはほぼ正常であるのに対して,さらにHes1遺伝子からイントロンを除去したところ,神経幹細胞の増殖・維持が障害されて,小頭症になることがわかった(図9C)25).したがって,Hes1の発現振動は,正しいタイミング(適切な時間遅れ)でネガティブ・フィードバックが起こることに依存し,正常な脳形成にきわめて重要であることが示された.
2)適切な時間遅れの細胞間カップリングに依存した発現振動
Hes1の発現は,隣接細胞のDll1によってNotchシグナルが活性化されることによって誘導される.神経発生過程では,NotchシグナルのリガンドDll1はプロニューラル因子によって活性化されてニューロンやニューロン前駆細胞に高レベルに発現する.しかし,ニューロンやニューロン前駆細胞が形成される前,すなわち,まだ神経幹細胞しか存在しないような神経発生の初期には,神経幹細胞自身がDll1を発現する.Dll1の発現はプロニューラル因子によって活性化され,Hes1によって抑制されるが,プロニューラル因子やHes1の発現は振動するのでDll1の発現も振動する15, 22, 27).Hes7のときと同様に,数理モデルの解析から,神経幹細胞どうしでNotchシグナルの活性化を行っている時期に,Dll1の発現のタイミングが加速あるいは遅延すると,Hes1の発現振動が減弱,あるいは定常発現になると予測された(図4C).そこで,上記のDll1の発現のタイミングを加速化した変異(Dll1 type-1変異)および遅延化した変異(Dll1 type-2変異)の2種類のDll1遺伝子ノックインマウスを解析したところ,数理モデルの予測どおり,Hes1の発現振動が減弱していた(図5A, C)15).さらに,神経幹細胞の増殖能が低下し,小頭症になった(図5C).したがって,Dll1の発現が正しいタイミングで起こること(適切な時間遅れの細胞間カップリング)がダイナミックな発現振動に必須であり,発現振動は神経幹細胞の増殖・維持に重要であることが明らかになった.
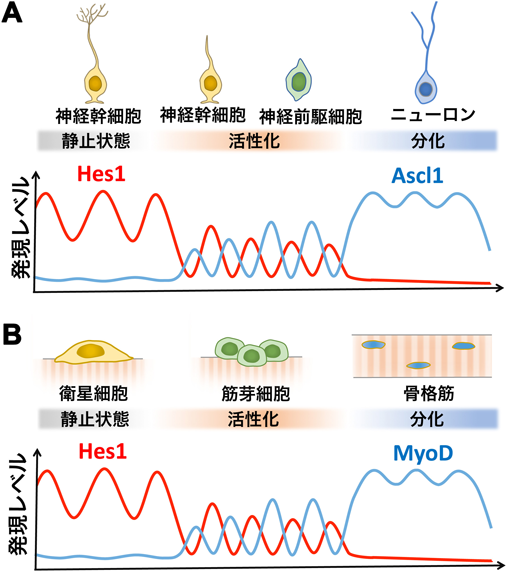
3)静止状態と活性化状態を制御するHes1とAscl1の発現動態
胎生期の神経幹細胞は,盛んに増殖しつつ多様な細胞を生み出す活性化状態にある.一方,成体脳の神経幹細胞は,ほとんど増殖しない静止状態にある28–32).この二つの状態の制御機構については不明の点が多いが,最近,Hes1やAscl1の発現動態が成体脳におけるニューロン新生でも重要な働きを持つことが明らかになってきた.上述のように,活性化状態の神経幹細胞では,Hes1の発現は振動し,Hes1によって周期的に抑制されるためにAscl1の発現も振動する22, 23).特に,Ascl1の発現が振動すると神経幹細胞の増殖が活性化され,Ascl1の発現がなくなると増殖能は低くなる23).成体脳の神経幹細胞も時折活性化して新たなニューロンを生み出すが,このときAscl1が必須な役割を担うことが報告されていた33, 34).
そこで,ライブ・イメージングによって成体脳の静止状態の神経幹細胞のHes1やAscl1の発現を解析したところ,Hes1の発現が増加し,高止まりで振動していた33).振動の谷でも十分に下がらず,活性化状態のときのHes1振動のピーク値レベルを維持していた.すなわち,静止状態の神経幹細胞ではHes1の発現は持続しており,その結果,Ascl1の発現はほぼ完全に抑制されていた.成体になってからHes1や関連遺伝子をノックアウトしたマウスでは,多くの神経幹細胞でAscl1の発現が起こり,数週間以内にほぼすべての神経幹細胞はニューロンに分化して枯渇した35).したがって,成体脳の神経幹細胞ではHes1が高レベルで持続発現することによってAscl1の発現が抑制され,その結果,静止状態になると考えられた(図10上).
それでは,静止状態の神経幹細胞にAscl1の発現振動を誘導すれば,活性化できるのだろうか? この問いに答えるために,光遺伝学的にAscl1を誘導する遺伝子セットをレンチウイルスによって成体脳の神経幹細胞に導入し,光ファイバーを挿入してパルス状に光照射を行った.1週間後に脳切片を作製して解析したところ,成体脳の神経幹細胞が効率よく活性化されて,幼若なニューロンを生み出していた35).また,神経幹細胞で発現振動を誘導できるHes5プロモーター下にAscl1をつないでレンチウイルスによって成体脳の神経幹細胞に導入したところ,光遺伝学的手法と同様に神経幹細胞が効率よく活性化されて,幼若なニューロンを生み出していた35).以上から,神経幹細胞は,Hes1の発現振動によってAscl1も発現振動すると活性化状態に,Hes1が高レベルで持続発現してAscl1の発現が抑制されると静止状態になることが示された(図10上).このAscl1の発現振動を誘導する手法は,成体脳に内在する神経幹細胞を再活性化して,損なわれた神経組織を再生する技術に応用されることが期待される.
他の組織形成においても遺伝子発現動態の重要性が明らかになってきた.骨格筋に存在する衛星細胞は骨格筋幹細胞で,普段は静止状態にある.しかし,筋肉が障害を受けたときなどに活性化されて,増殖を開始し,ある程度細胞数が増えてから筋肉に分化する.ライブ・イメージングの解析から,静止状態の衛星細胞ではHes1が高レベルで持続発現して筋肉分化決定因子MyoDの発現が抑制されていた.しかし,活性化状態(筋芽細胞)になるとHes1の発現振動によってMyoDも発現振動することがわかった(図10下)36).ある程度増殖した後,Hes1の発現はなくなり,MyoDは持続発現して筋肉分化が起こった.さらに,このMyoDの発現振動が骨格筋幹細胞(筋芽細胞)の増殖に重要な役割を担うことが示された36, 37).したがって,骨格筋幹細胞の静止状態,活性化状態,筋肉分化過程のいずれもがHes1とMyoDとの発現動態の変化に深く依存することが明らかになった(図10下).
同様に,膵形成においてもHes1の発現振動によって組織幹細胞の増殖が活性化されており,Hes1の発現振動が減衰すると膵臓は低形成に陥ることがわかった38).Hes1の発現振動はその他の多くの細胞でもみられることから,多くの組織形成に共通な役割を担うと考えられた.