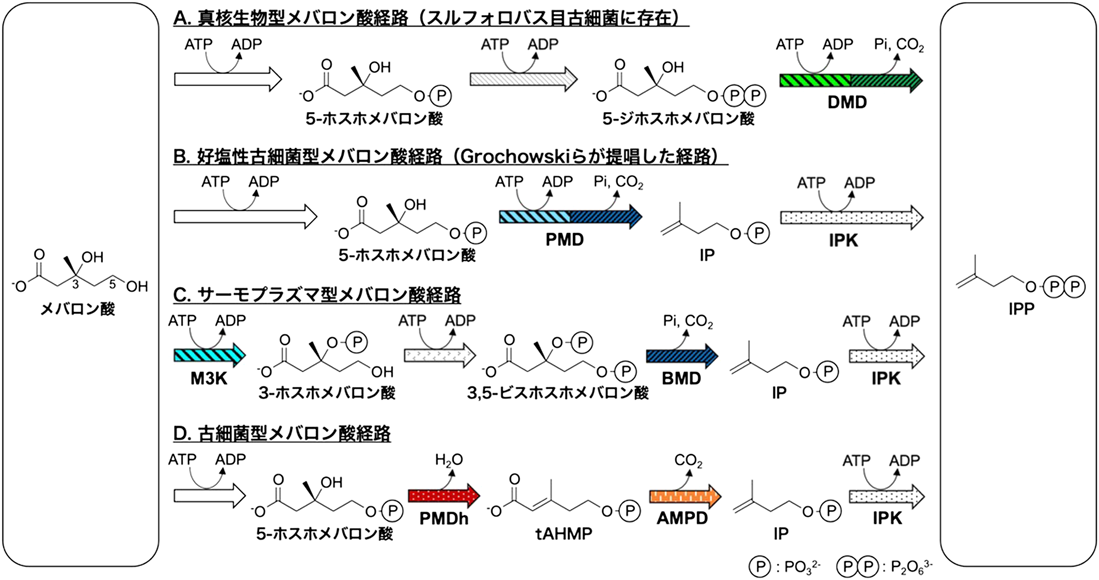
古細菌型メバロン酸経路の発見Discovery of the archaeal mevalonate pathway
名古屋大学大学院生命農学研究科Graduate School of Bioagricultural Sciences, Nagoya University ◇ 〒464–8601 愛知県名古屋市千種区不老町 ◇ Furo-cho, Chikusa-ku, Nagoya, Aichi 464–8601, Japan
発行日:2021年4月25日Published: April 25, 2021