我々現在の地球生命は,多様であると同時に,なぜ普遍的に20種類のアミノ酸を共通してリボソームにおけるタンパク質合成に用いているのだろうか.博物学として古代から進展してきた生物学について,前世紀の生化学・分子生物学の展開から共通性が見いだされたことに非常な興奮があったことは間違いないだろう.だからこそ,中心教義(セントラルドグマ)や,普遍遺伝暗号(universal genetic code),という神や宇宙と関連する単語が用いられたのではなかろうか.その後の諸研究により,遺伝暗号における20種類のアミノ酸の配置が変化した事例や,アミノ酸の追加事例としてmRNAの文脈に依存して導入されるセレノシステイン,一部の微生物にみられるピロリシンといった事例が見つかってきたものの,リボソームにおけるタンパク質合成がこれら22アミノ酸によって寡占されている.
一方,生命における普遍性を議論する際に,現在の地球生命は生命の一つのかたちにすぎない,ということに注意する必要がある.すなわち,我々が自然界から見いだす共通性が,この宇宙の物理化学法則によって普遍性に制約されているものなのか,46億年間の地球環境に制約されているものなのか,進化的に偶然凍結されたものか,といった視点が重要になる1).
生物があるアミノ酸を活用するか否かは,そのアミノ酸が外界または自らによる生合成で供給されるか,そのアミノ酸を重合装置で利用可能であるか,によって決められている.さらには,ひとたび使用され出すと,そのアミノ酸を生命システムから追い出すことができない,という意味で,「利己的なアミノ酸」となったものがあるかもしれない2).
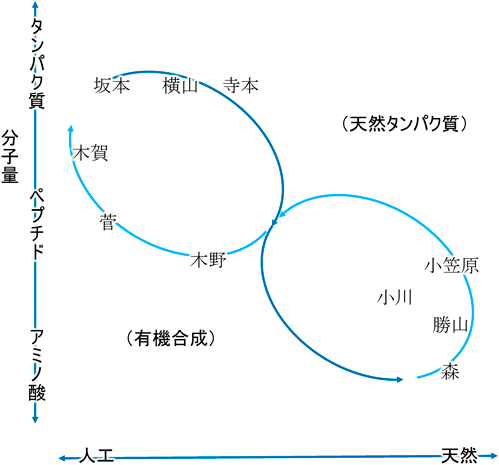
本特集では,これら22種類以外,すなわち,通常ではリボソームではタンパク質に取り込まれない種々のアミノ酸に注目し,これらについての議論を深めるために,重合系,モノマー供給系に関する執筆を各分野の第一人者の先生方にお願いした(図1).また,天然における現象の発見だけでなく,共通性を突破するための種々の改変についても網羅している.新しい現象は,天然由来のもの,人工的なもの,どちらであっても工学的なツールとなり,種々の非標準アミノ酸の合成ならびにこれらを取り込んだペプチド・タンパク質の有効活用事例が蓄積されている.非標準アミノ酸のうち,天然でのリボソームにおける取り込みがみられるセレノシステイン,ピロリシンに関しては,本会誌の過去の記事等を参照されたい3–6).
坂本の記事が,宮澤博士が主導した「超タンパク質」の紹介から始まり,アミノアシルtRNA合成酵素(aaRS)変異体−tRNA変異体を用いることで標準遺伝暗号を拡張して遺伝暗号に新たなアミノ酸を加える方法について詳述するとともに,横山の稿では,「生きるための制約を外す」ことが可能な無細胞系における遺伝暗号の拡張について詳述する.遺伝暗号内のアミノ酸セットの変更は動物体内でも可能であり,aaRS変異体を介してカイコ体内でフェニルアラニン類縁体を組み込んだシルクタンパク質を生産する研究が寺本の稿で記されている.
リボソーム上でのタンパク質合成よりも広く視点をとると,より多様なアミノ酸が自然界で生合成されている.森,勝山,小川の各稿は,これらのアミノ酸モノマーの生合成過程を紹介する.森は植物における非タンパク質性のアミノ酸の意義と合成について詳述する.勝山の記事では,放線菌での非リボソームペプチド合成に供給される非標準アミノ酸の生合成を紹介する.小川は,アミノ酸水酸化酵素による自然界での非標準アミノ酸の同化・異化のみならず,この酵素を活用した工学アプローチについて紹介する.その中で,隕石中やミラーの化学進化実験でみられる一方,天然にはみられないアミノ酸であるAibを基質とした反応が興味深い.
小笠原,木野,菅による記事はペプチド合成を扱う.小笠原は,リボソーム,非リボソーム,それぞれで生合成されるペプチドにおけるD-アミノ酸に焦点をあてた,自然界が生み出した多様な合成系について解説を記している.木野は,酵素を用いたジペプチドの合成に種々の非標準アミノ酸も活用することが可能であり,生成物が医薬品原料として期待されていることを紹介する.菅らの合成系はリボソームを活用するものであるが,リボザイムにより多様な非標準アミノ酸をtRNAに結合させることを活かした,創薬を志向した特殊環状ペプチドの創出が可能になっていることを詳述する.
これら各稿は,通常ではリボソームにおけるタンパク質合成には用いられない多様なアミノ酸が酵素によって合成されることが可能であり,また,酵素によってこれら非標準アミノ酸がペプチドやタンパク質に導入可能なことを示している.その意味から,現在の20種類のアミノ酸を用いた標準遺伝暗号は,「ありえた標準」の一つでしかないといえよう.この観点から,過去にありえた,また,別の進化として生じえた遺伝暗号について私の記事を最後に加えた.
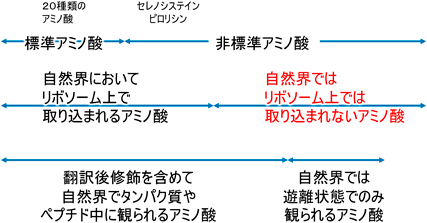
なお,「普遍」遺伝暗号表に記された20種類以外のアミノ酸について,種々の区分が存在する(図2).まず,遺伝暗号について,「普遍」遺伝暗号以外の暗号が種々の生物で見つかっており普遍性が弱まったことから,「標準遺伝暗号」と呼ばれることが多い.標準遺伝暗号に記された,という意味で,20種類のアミノ酸は標準アミノ酸と呼ぶことができよう.これら以外を非標準アミノ酸と呼ぶと,そのうちのセレノシステインとピロリシンはリボソームでタンパク質に取り込まれる.一方,遺伝暗号の拡張によって天然においてはリボソームでの翻訳では使用されないアミノ酸を翻訳系で使用可能とする際に,「非天然型」アミノ酸という呼称が使用されることもある.この区分のアミノ酸について,「非翻訳性アミノ酸」「非新生鎖性アミノ酸」と呼ぶこともできるかもしれないが,異論も多いだろう.また,ヨードチロシンなど,タンパク質の翻訳後修飾によってタンパク質中に存在するアミノ酸残基もこの定義に含まれることに留意されたい.関連して,タンパク質性アミノ酸,という用語にも注意が必要である.Proteinogenic amino acidには,20種類の標準アミノ酸に加え,セレノシステイン,ピロリシン,場合によってはホルミルメチオニンが含まれる.一方,日本語として「タンパク質性」と呼ぶ場合には,新生鎖のみ,もしくは,翻訳後修飾を含めてタンパク質ととらえるかで,意味合いが変わりうる.本特集を通じてお示しできることは,天然でのアミノ酸の合成系と重合系の多様性である.おそらく,天然には未知のものが存在し,将来の発見によって,アミノ酸セットの呼称と内容が再考されるであろう.
謝辞Acknowledgments
末筆ながら,本企画に御賛同下さり,皆様の多くの研究の中から本企画に焦点を当てた執筆をしていただいた各先生に深謝申し上げます.
引用文献References
1) クリスティアン・ド・デューブ著,中村桂子監訳(2007)進化の特異事象,一灯舎.
2) Amikura, K. & Kiga, D. (2013) The number of amino acids in a genetic code. RSC Advances, 3, 12512–12517.
3) 松儀実広(2018)セレノプロテイン生合成系について.生化学,79, 964–968.
4) Itoh, Y., Bröcker, M.J., Sekine, S., Hammond, G., Suetsugu, S., Söll, D., & Yokoyama, S. (2013) Decameric SelA·tRNA(Sec) ring structure reveals mechanism of bacterial selenocysteine formation. Science, 340, 75–78.
5) Tsukazaki, T., Mori, H., Fukai, S., Ishitani, R., Mori, T., Dohmae, N., Perederina, A., Sugita, Y., Vassylyev, D., Ito, K., et al. (2008) Conformational transition of Sec machinery inferred from bacterial SecYE structures. Nature, 455, 988–991.
6) 野澤佳世,石谷隆一郎,濡木 理(2010)ピロリジン翻訳の直交性のメカニズム.生化学,82, 617–623.