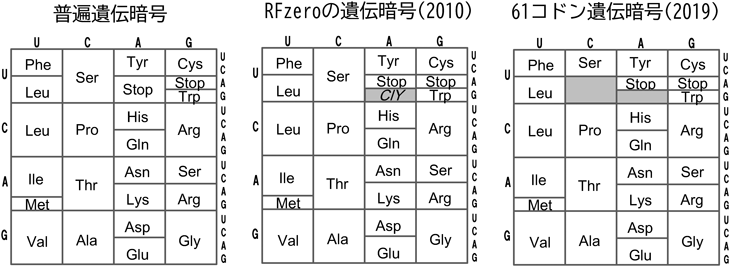
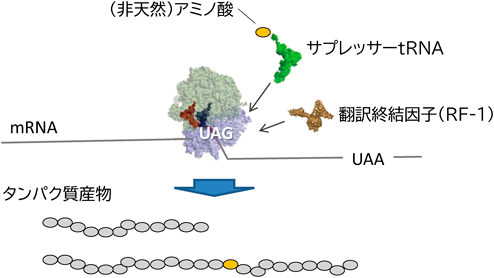
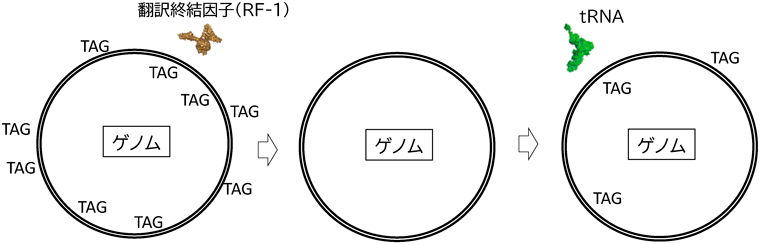
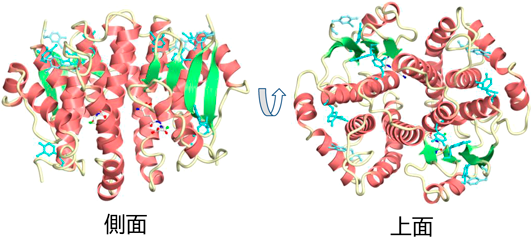
生細胞における遺伝暗号の改変と応用In vivo engineering of the genetic code and its applications
理化学研究所・生命機能科学研究センターRIKEN Center for Biosystems Dynamics Research ◇ 〒230–0045 神奈川県横浜市鶴見区末広町1–7–22 ◇ 1–7–22 Suehiro-cho, Tsurumi-ku, Yokohama, Kanagawa 230–0045, Japan
発行日:2021年6月25日Published: June 25, 2021