カイコ(蚕,Bombyx mori)は鱗翅目(Lepidoptera)に属する昆虫の1種であり,「繊維の女王」とも形容されるシルク(絹)を作る虫であることは広く知られている.日本では明治から昭和中ごろにかけて養蚕が盛んに行われ,1929年(昭和4年)には全国の養蚕農家戸数が221万戸にも達した1).その後はシルク需要や価格の低迷により戸数は減少を続け,2019年には全国で264戸を残すのみとなった2).現代に生きる日本人にとって養蚕は縁遠いものとなっている.
シルクを身につける利点としてよくあげられるのは,その着心地の良さである.シルクはタンパク質でできているため肌への刺激が少ないといわれる.手術用の縫合糸としても長年使われており,強度に優れ生体適合性が高い素材であることがわかっている3).絶縁性にも優れ,電子材料としての利用研究も進んでいる4).人類がシルクを発見してから数千年を経てもなお,そのユニークな特性は研究者を惹きつけている.
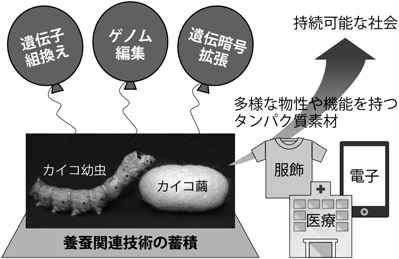
産業としての養蚕は国内では衰退してしまったものの,カイコを飼育して繭を収穫しシルクを取り出す一連の高度な技術は蓄積されている.また鱗翅目を代表するモデル生物としてのカイコの基礎研究が盛んに行われ,全ゲノム配列5),遺伝子組換え6),ゲノム編集7)といったバイオテクノロジーに関わる情報や技術が整備されている.そこで,伝統的な養蚕技術と最新のバイオテクノロジーとを組み合わせる新たな形での養蚕業の再開拓が期待されている.
本稿では,カイコの新たなバイオテクノロジーとして遺伝暗号拡張(翻訳システムの改変)の実現に向けた取り組みを取り上げ,その現状を紹介する.カイコで遺伝暗号拡張を実現すれば,非天然アミノ酸を含有するタンパク質を大量生産し,繊維等の素材として利用することができるようになる.さらに遺伝子組換えやゲノム編集と組み合わせることによって,多様な物性や機能を持つタンパク質素材が創製できる.折しもプラスチックによる環境汚染が大きな社会問題となり,脱プラスチックが盛んに叫ばれている.天然のタンパク質を基盤としつつバイオテクノロジーによって物性や機能を最適化した素材は,服飾・医療・電子等の幅広い分野での活用を通して持続可能な社会の構築に貢献できるだろう.
カイコは卵からふ化して繭を作り始めるまでの約1か月の間に体重が1万倍にも増加し,1頭が0.3グラム程度のシルクを繊維として紡ぎ出す8).大腸菌に0.3グラムの組換えタンパク質を作らせる大変さを考えれば,カイコのタンパク質合成能力は驚異的である.特にシルクのような高分子量かつ反復配列に富んだ繊維タンパク質の大量生産は,大腸菌や培養細胞での発現や化学合成法では実現が困難である.このカイコの優れたタンパク質合成能力はこれまでシルク生産のみに利用されてきたわけだが,2000年に遺伝子組換え技術が確立されたことによってシルク以外の外来タンパク質や,機能性タンパク質を融合した改変シルクをカイコに作らせることが可能となった9, 10).さらに近年ではゲノム編集によってシルクの遺伝子を丸ごと外来遺伝子に置き換え,より効率的に外来タンパク質を作らせることもできるようになってきている11).
図1に,我々の研究が目指す方向性をまとめる.カイコという生物自体が持つ能力,すなわち,繊維タンパク質を大量に合成し繊維という素材形態で分泌できる能力は,他の生物と比較して非常にユニークといえる.さらにバイオテクノロジーを融合させることで,カイコのもつ能力を人類にとってより有効に活用することが可能である.カイコは繊維形態でタンパク質を分泌するが,水溶液やフィルム,ゲル,ナノファイバー,ナノ粒子といった異なる素材形態へと加工することも容易である12).生産性やコストといった課題はもちろんあるだろうが,環境や健康への影響を低減させるための一つのアプローチとして有望と考えている.
20種類の標準アミノ酸にセレノシステインとピロリシンとを加えた22種類以外のアミノ酸は,非天然アミノ酸(あるいは人工アミノ酸)と呼ばれる.非天然アミノ酸は任意の官能基や分子団を導入したものが設計できるため,天然にはない物性や機能を持つペプチドやタンパク質の合成に利用できる.当然ながら,通常は生物がこのような非天然アミノ酸をタンパク質合成に利用することはできない.それを実現する方法論が遺伝暗号拡張(genetic code expansion)である13, 14).
遺伝暗号拡張とは,生物の遺伝暗号にコードされるアミノ酸の種類を増やすための方法論のことをいう.遺伝暗号はmRNA上の各コドンをどのアミノ酸として翻訳するかという対応関係を指定している.コドンとアミノ酸との対応関係の正確さは,主としてアミノアシルtRNA合成酵素(aminoacyl-tRNA synthetase:aaRS)の働きによって担保されている.aaRSはtRNAの3´末端にアミノ酸を結合させる酵素であり,アミノ酸ごとに専用のaaRSが存在する.aaRSがアミノ酸とtRNAとの正しい組合わせを選び出すことによってmRNA上のコドンが正しいアミノ酸として翻訳されることになる.
遺伝暗号拡張を実現するための比較的簡便な手法として,非天然アミノ酸を基質として認識するように改変したaaRS変異体を発現させるという手法がある.このようなaaRS変異体を発現する生物は,標準アミノ酸に加えて非天然アミノ酸もタンパク質の合成に用いることができるようになる.たとえば,カリフォルニア工科大学のTirrellらのグループは,非天然アミノ酸(4-bromophenylalanine:BrPhe)を認識するように改変したフェニルアラニルtRNA合成酵素(PheRS)変異体を発現させた大腸菌をBrPheの存在下で培養することによって,タンパク質(dihydrofolate reductase:DHFR)中にBrPheが導入されることを2000年に報告した15).この手法では,フェニルアラニン(Phe)を指定するコドン(UUUおよびUUC)がPheあるいはBrPheのいずれかとして翻訳されることになる.Tirrellらは上記の論文で,培地からPheを除去することによってDHFR中のPheコドン(9個)のうちの約9割がBrPheとして翻訳されることを示した15).
上記の手法は,もともと標準アミノ酸に指定されているコドンをそのまま利用し,標準アミノ酸として翻訳されるはずのコドンを非天然アミノ酸として翻訳させる手法である.したがって,細胞内に標準アミノ酸が一切存在しない理想的な条件下では,あるアミノ酸残基(上述の例ではPhe)を指定するすべてのコドンに対して非天然アミノ酸が導入されることになる.このような遺伝暗号拡張手法は「残基特異的」手法と呼ばれ,複数の非天然アミノ酸をタンパク質中に導入したい場合に有効である.この手法でaaRSを改変せずともメチオニンの代わりにノルロイシンを組み込んだ「超タンパク質」を大腸菌に生産させた例16)に対して,本研究ではaaRSの改変によって初めて取り込みが可能になった非天然アミノ酸を活用している.一方,本手法ではタンパク質中での非天然アミノ酸の導入位置を厳密に指定することはできない.
遺伝暗号拡張の他の手法については,本特集の他編(坂本,横山,菅の各編)あるいは上にあげた参考文献13, 14)をご参照いただきたい.
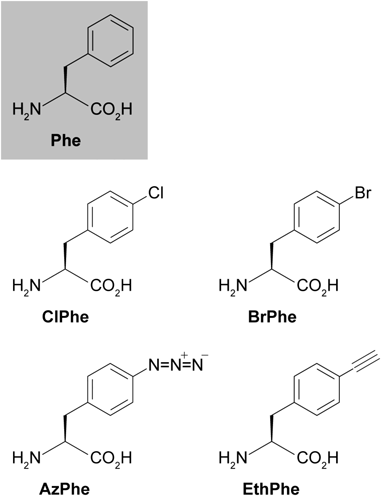
筆者らはカイコでの遺伝暗号拡張に挑戦するにあたり,残基特異的手法を用いることにした.その理由として,①aaRS変異体のみを発現させるだけで実現可能なシンプルな手法であること,②一つのタンパク質分子内に複数の非天然アミノ酸を導入するのに適していること,の2点があげられる.どのアミノ酸残基をターゲットとするかについては,上述したTirrellらの先行研究と同じくPheを選んだ.シルク中でのPheの存在量は約0.6%と少なくシルクの構造形成に大きく関与しないと考えられ,ターゲットとして好都合であった.またPheはカイコが生合成できない必須アミノ酸であり,飼料組成によって細胞内での存在量を制御できることも利点であった.
先行研究でTirrellらが用いていたPheRS変異体は大腸菌由来であり,カイコの細胞内ではうまく機能しないと予想された.そこでカイコのPheRSをクローニングし,大腸菌由来PheRS変異体と相同の変異を導入したカイコPheRS変異体3種を作出した.これらの変異体はPheに構造が類似した非天然アミノ酸(Phe類縁体)を認識できることをin vitro実験およびカイコ培養細胞を用いた実験で確認した17–19).そこでこれらのPheRS変異体を絹糸腺(シルク合成器官)で発現する遺伝子組換え(TG)カイコを作出した.Phe類縁体を混合した飼料をTGカイコに摂食させ,分泌されたシルクを解析したところ,シルク中のPheの一部がPhe類縁体に置き換わっていることを確認した20–22).これらの実験により,カイコにも遺伝暗号拡張の手法が適用可能であり,かつ,非天然アミノ酸を含有するシルクを作出できることが示された(図2).本手法により,これまでに4種類のPhe類縁体(図3)がシルクに導入可能であることを確認している.
4. クリック反応で簡単に機能付加できる「クリッカブルシルク」の開発
図3に示したPhe類縁体のうち,特にAzPheに着目した.AzPheに含まれるアジド基は天然の生体分子中には存在せず,アルキン基と選択的に反応するという性質を持つ.このアジド基とアルキン基との間での反応(アジド-アルキン付加環化反応)はクリックケミストリー(click chemistry)の代名詞的反応としてよく知られ,化学,生物学,薬学,材料科学に至る幅広い分野で多くのブレイクスルーをもたらしている23).本稿ではアジド-アルキン付加環化反応のことを単に「クリック(click)反応」と呼ぶ.AzPheを導入したシルクでは,クリック反応によってさまざまな機能分子を結合させることができる.このような技術は,服飾・医療・電子等への応用で求められるさまざまな性質を柔軟にシルクに付加する基盤技術として汎用性が高い.筆者らはAzPheを導入したシルクのことを「クリッカブルシルク(clickable silk)」と呼び,その社会実装に向けた取り組みを進めている.
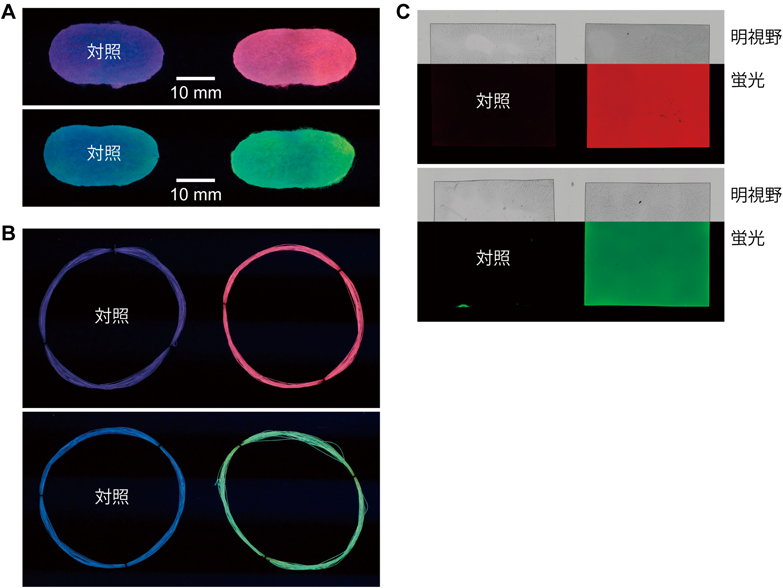
開発初期のTGカイコ系統では,シルクへのAzPheの導入効率が低いことが課題であった.そこで筆者らは理化学研究所の坂本健作博士らと共同で,AzPheの導入効率を大幅に向上させたTGカイコ系統(H06系統)を作出した24).H06系統は,新たに見いだしたカイコPheRS-F432V変異体を絹糸腺で発現している.H06系統を用いると,全Phe残基の約7%がAzPheに置き換わったシルクを生産することができる.これはシルク分子一つあたりにおよそ2.5個のAzPheが導入されていることに相当する.H06系統にAzPheを投与して作らせた繭にアルキン基を持つ蛍光分子をクリック反応させたところ,鮮やかな蛍光色を示した(図4A).その他,繊維やフィルム等の他の形状に加工した後でもクリック反応が問題なく進行し,明瞭な蛍光が観察された(図4B, C).
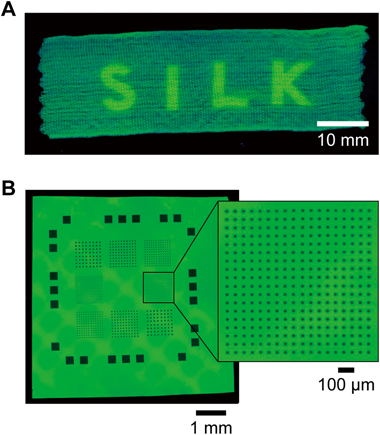
AzPhe中のアジド基のように芳香環に直接結合したアジド基(芳香族アジド)は紫外線照射によって分解することが知られている25, 26).この性質を利用すれば,たとえば布やフィルムといった平面シルク上でアジド基の存在量に濃淡をつけることができると考えた.実際に布を作り,「SILK」の文字シールを貼って紫外線を照射したのち蛍光色素とのクリック反応を行ったところ,SILKの文字パターンが明瞭に現れた(図5A).同様の方法を用いてフィルム上にμmスケールの微細なパターンを作ることもできた(図5B).この方法を利用すればシルクの性質をμmスケールで精密に制御できると考えられる.
本稿の2節に,カイコ1頭が0.3グラム程度のシルクを繊維として紡ぎ出すと書いた.ところが実際には,H06系統が作るシルクの量は約0.1グラムである.これはTGカイコの作出に実験が容易な小型の系統を用いたためである.社会実装に向けては1頭あたりのシルク生産量を少なくとも2倍以上に向上させる必要があると考えている.
シルク生産量を向上させる単純な方法として,たくさんのシルクを作る大型の系統と掛け合わせる方法がある.そこでH06系統を日509×日510という大型の野生型系統と掛け合わせた交雑種を作ったところ,そのシルク生産量はH06系統の約1.5倍に向上した27).野生型と交雑することでPheRS変異体の発現量が低下し,それに伴いAzPheの導入効率も低下する懸念があったが,実際に調べてみるとAzPheの導入効率に変化はなかった27).大型化した交雑種の繭からはシルク繊維を容易に調製できるようになり,布等の実用的な材料形態に加工するのに十分な量のクリッカブルシルク繊維を得ることができるようになった(図6).しかしながら,大型の系統との交雑だけでは目標とする2倍以上のシルク生産量に届かない.そこで現在,シルク生産量をさらに増大させるための系統改良を進めている.
最後に,クリッカブルシルクの応用例を一つ紹介したい.
シルクは縫合糸として使われるほど生体適合性が高いことは先に述べた3).繊維だけでなくフィルムなどさまざまな形状に加工しやすいことから,バイオマテリアルとしての応用研究が盛んに行われている.バイオマテリアルへの応用においては,基材表面と細胞との相互作用をいかに制御するかがとても重要となる.クリッカブルシルクを原料として用いれば,異なる表面性状を持つシルク基材をクリック反応によって調製できると考えられる.そこで,細胞接着を抑制することが知られているポリエチレングリコール(PEG)をクリック反応で結合させたシルクフィルムを調製し,マウス線維芽細胞(NIH3T3)を用いてフィルム表面への細胞接着数の変化を調べた.その結果,PEGを修飾したシルクフィルム表面への接着細胞数は予想どおり大きく減少した28).上述したように,シルクフィルム上でのアジド基の存在は紫外線照射によって制御できる.そこでクリッカブルシルクフィルムにあらかじめ紫外線を照射して部分的にアジド基を破壊し,残存したアジド基に対してクリック反応でPEGを結合させた.このようにして調製したフィルム上でNIH3T3細胞を培養したところ,フィルム上の紫外線照射部位(=PEGが結合していない部位)でのみ細胞を増殖させることができた(図7).この手法を用いればシルク基材上で細胞を狙いどおりに配列させたバイオマテリアルを創製できると考えられる.
本稿では,カイコにおける遺伝暗号拡張(翻訳システムの改変)の実現によるシルクへの非天然アミノ酸の導入と,クリック反応によって望みの機能を簡単に付加できるタンパク質素材「クリッカブルシルク」の開発について紹介した.シルクに導入可能な非天然アミノ酸の種類や導入量にはまだまだ制約があるものの,高機能なタンパク質素材を作り出す上で遺伝暗号拡張の手法が有効であることを示すことができた.大量飼育の技術が確立されているカイコを用いることで,非天然アミノ酸を含むタンパク質素材を容易に大量生産できることが大きな特徴である.今後は,生産コストを低減させる試みと同時に,どのような分野への応用が有効かを見きわめていく必要がある.民間企業を含めた外部機関との連携により基礎と応用の両面から社会実装に向けた取り組みを進めていきたい.
本稿で紹介したカイコの遺伝暗号拡張による非天然アミノ酸含有タンパク質の生産手法は,当然ながらシルク以外のタンパク質についても適用可能である.カイコは薬剤タンパク質をはじめとする機能性タンパク質の生産宿主としてもすでに実用化されている9, 29).遺伝暗号拡張の手法を取り入れることによってタンパク質生産宿主としてのカイコの魅力がさらに増すことを期待している.
伝統的な養蚕技術は日本が世界に誇る技術であるものの,養蚕業の衰退とともに失われようとしている.最新のバイオテクノロジーと融合させることによって日本発の新型養蚕業の開拓を実現したい.
謝辞Acknowledgments
本特集で執筆する機会を与えて下さいました早稲田大学先進理工学部木賀大介先生に厚く御礼申し上げます.本稿で紹介した研究成果は,理化学研究所坂本健作博士,信州大学繊維学部玉田靖教授,農研機構生物機能利用研究部門小島桂博士らをはじめとする多くの共同研究者の皆様の協力によって得られたものです.この場をお借りして御礼申し上げます.なお本稿で紹介した研究の大部分は科研費(15K07800, 24658056, 24688008, 22658019, 20780039)の支援により行われたものです.研究を進めるにあたって多大な協力をいただいた研究支援者の皆様に感謝します.
引用文献References
1) 農林水産省(2020)蚕糸業をめぐる事情(https://www.maff.go.jp/j/seisan/tokusan/attach/pdf/sannshi-1.pdf).
2) (一財)大日本蚕糸会(2020)養蚕農家数の推移.シルクレポート,67, 39.
3) Holland, C., Numata, K., Rnjak-Kovacina, J., & Seib, F.P. (2018) The biomedical use of silk: Past, present, future. Adv. Healthc. Mater., 8, e1800465.
4) Wang, C., Xia, K., Zhang, Y., & Kaplan, D.L. (2019) Silk-based advanced materials for soft electronics. Acc. Chem. Res., 52, 2916–2927.
5) International Silkworm Genome Consortium. (2008) The genome of a lepidopteran model insect, the silkworm Bombyx mori. Insect Biochem. Mol. Biol., 38, 1036–1045.
6) Tamura, T., Thibert, C., Royer, C., Kanda, T., Abraham, E., Kamba, M., Komoto, N., Thomas, J.L., Mauchamp, B., Chavancy, G., et al. (2000) Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotechnol., 18, 81–84.
7) Ma, S.-Y., Smagghe, G., & Xia, Q.-Y. (2019) Genome editing in Bombyx mori: New opportunities for silkworm functional genomics and the sericulture industry. Insect Sci., 26, 964–972.
8) 農業・食品産業技術総合研究機構(2020)カイコってすごい虫!(http://www.naro.affrc.go.jp/nias/gmo/files/kaikottesugoimushi2020.pdf).
9) Xu, H. (2014) The advances and perspectives of recombinant protein production in the silk gland of silkworm Bombyx mori. Transgenic Res., 23, 697–706.
10) 小島 桂(2016)遺伝子組換えカイコが開くシルク利用の最前線.化学と生物,54, 915–919.
11) Xu, J., Dong, Q., Yu, Y., Niu, B., Ji, D., Li, M., Huang, Y., Chen, X., & Tan, A. (2018) Mass spider silk production through targeted gene replacement in Bombyx mori. Proc. Natl. Acad. Sci. USA, 115, 8757–8762.
12) Rockwood, D.N., Preda, R.C., Yücel, T., Wang, X., Lovett, M.L., & Kaplan, D.L. (2011) Materials fabrication from Bombyx mori silk fibroin. Nat. Protoc., 6, 1612–1631.
13) Mukai, T., Lajoie, M.J., Englert, M., & Söll, D. (2017) Rewriting the genetic code. Annu. Rev. Microbiol., 71, 557–577.
14) de la Torre, D. & Chin, J.W. (2020) Reprogramming the genetic code. Nat. Rev. Genet., 22, 169–184.
15) Sharma, N., Furter, R., Kast, P., & Tirrell, D.A. (2000) Efficient introduction of aryl bromide functionality into proteins in vivo. FEBS Lett., 467, 37–40.
16) Koide, H., Yokoyama, S., Kawai, G., Ha, J.M., Oka, T., Kawai, S., Miyake, T., Fuwa, T., & Miyazawa, T. (1988) Biosynthesis of a protein containing a nonprotein amino acid by Escherichia coli: L-2-aminohexanoic acid at position 21 in human epidermal growth factor. Proc. Natl. Acad. Sci. USA, 85, 6237–6241.
17) Teramoto, H. & Kojima, K. (2010) Cloning of Bombyx mori phenylalanyl-tRNA synthetase and the generation of its mutant with relaxed amino acid specificity. J. Insect Biotechnol. Sericol., 79, 53–65.
18) Teramoto, H. & Kojima, K. (2013) Residue-specific incorporation of phenylalanine analogues into protein biosynthesis in silkworm cultured cells. J. Insect Biotechnol. Sericol., 82, 61–69.
19) Teramoto, H., Kojima, K., Kajiwara, H., & Ishibashi, J. (2012) Expansion of the amino acid repertoire in protein biosynthesis in silkworm cells. ChemBioChem, 13, 61–65.
20) Teramoto, H. & Kojima, K. (2014) Production of Bombyx mori silk fibroin incorporated with unnatural amino acids. Biomacromolecules, 15, 2682–2690.
21) Teramoto, H., Nakajima, K., & Kojima, K. (2016) Azide-incorporated clickable silk fibroin materials with the ability to photopattern. ACS Biomater. Sci. Eng., 2, 251–258.
22) Teramoto, H. (2017) In vivo incorporation of an alkyne-bearing amino acid into Bombyx mori silk fibroin. J. Insect Biotechnol. Sericol., 86, 113–121.
23) Xi, W., Scott, T.F., Kloxin, C.J., & Bowman, C.N. (2014) Click chemistry in materials science. Adv. Funct. Mater., 24, 2572–2590.
24) Teramoto, H., Amano, Y., Iraha, F., Kojima, K., Ito, T., & Sakamoto, K. (2018) Genetic code expansion of the silkworm Bombyx mori to functionalize silk fiber. ACS Synth. Biol., 7, 801–806.
25) Reddington, S., Watson, P., Rizkallah, P., Tippmann, E., & Jones, D.D. (2013) Genetically encoding phenyl azide chemistry: New uses and ideas for classical biochemistry. Biochem. Soc. Trans., 41, 1177–1182.
26) Tippmann, E.M., Culpepper, S., Bunnel, W., & Appel, N. (2019) New perspectives on aryl azide noncanonical amino acid use in yeast. Photochem. Photobiol. Sci., 18, 253–258.
27) Teramoto, H., Iga, M., Tsuboi, H., & Nakajima, K. (2019) Characterization and scaled-up production of azido-functionalized silk fiber produced by transgenic silkworms with an expanded genetic code. Int. J. Mol. Sci., 20, 616.
28) Teramoto, H., Shirakawa, M., & Tamada, Y. (2020) Click decoration of Bombyx mori silk fibroin for cell adhesion control. Molecules, 25, 4106.
29) Itoh, K., Kobayashi, I., Nishioka, S., Sezutsu, H., Machii, H., & Tamura, T. (2016) Recent progress in development of transgenic silkworms overexpressing recombinant human proteins with therapeutic potential in silk glands. Drug Discov. Ther., 10, 34–39.
著者紹介Author Profile
 寺本 英敏(てらもと ひでとし)
寺本 英敏(てらもと ひでとし)国立研究開発法人農業・食品産業技術総合研究機構生物機能利用研究部門グループ長補佐.博士(工学).
略歴1975年兵庫県生まれ.98年大阪市立大学理学部物質科学科卒業.2000年同大学院物質分子系専攻修士課程修了.同年農林水産省蚕糸・昆虫農業技術研究所入所.07年東京農工大学大学院工学府生命工学専攻博士課程修了.16年より現職.
研究テーマと抱負天然のシルクをベースに高機能な素材を作り出すための技術開発を進めています.天然素材を高度に活用する技術を開発することで持続可能な社会の構築に少しでも貢献できたらと思っています.
ウェブサイトhttp://www.naro.affrc.go.jp/laboratory/nias/introduction/chart/0204/index.html
趣味テニス,映画鑑賞,旅行.