微生物や植物はその生命活動のためさまざまなペプチド化合物を利用する.それら化学構造は多様であり,骨格に非タンパク質性アミノ酸やD-アミノ酸を含有するものなどが数多く存在する.細菌の代表的なD-アミノ酸含有ペプチドとして細胞壁の構成成分であるペプチドグリカンがあげられ,ペプチド鎖にD-GluやD-Alaを含みプロテアーゼによる加水分解を受けにくい構造になっている1).また,ペニシリンやバンコマイシンなどの二次代謝産物にもD-アミノ酸が含まれており,他の非タンパク質性アミノ酸とともにペプチド構造の多様性の拡大や生理活性の発現に寄与している2).
D-アミノ酸含有ペプチドの生合成では,遊離のD-アミノ酸の生成後にアミノ酸リガーゼによりペプチド結合が形成される場合と,ペプチド結合形成後にエピメリ化が起こる場合がある.前者の場合,遊離のD-アミノ酸は主にアミノ酸ラセマーゼによって生合成される.細菌はさまざまなアミノ酸に対応する複数のアミノ酸ラセマーゼを有するが,ピリドキサール5′-リン酸(pyridoxal 5′-phosphate:PLP)を補酵素に用いるPLP依存酵素とPLP非依存酵素の二つのタイプがある.また,D-アミノ酸アミノ基転移酵素によりα-ケト酸から生合成される経路も知られている.これらの遊離D-アミノ酸の生合成についての詳細は,他の総説に詳しく記載されているのでそちらを参照いただきたい3–5).これに対して,後者のペプチドを基質とするエピメリ化反応が起こる場合も,一次代謝のペプチドグリカンの分解経路や一部の微生物のみが利用するペプチドグリカン生合成経路に見いだされている.また,二次代謝産物の生合成でもペプチドを基質とするエピメリ化反応でD-アミノ酸残基を導入する例がいくつか知られている.本稿では,それらの詳細について著者らの研究成果を含めて紹介する.
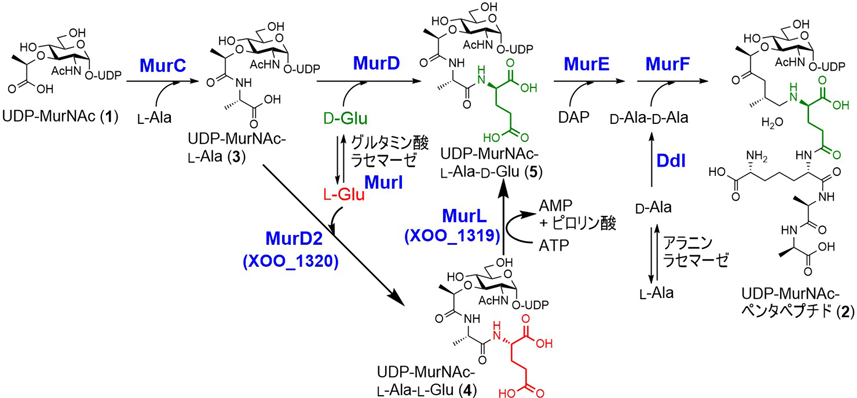
ほぼすべての細菌は,細胞の構造維持のため細胞壁にペプチドグリカンを有する.ペプチドグリカンは,N-アセチルグルコサミン(N-acetylglucosamine:GlcNAc)とN-アセチルムラミン酸(N-acetylmuramic acid:MurNAc)の繰り返しで構成される糖鎖のMurNAcにオリゴペプチドが結合し,さらにペプチド鎖どうしがクロスリンクすることで強固な網目構造を形成している1).ペプチド鎖にはD-GluやD-Alaが含まれるが,通常の生合成では,生合成中間体であるウリジン二リン酸(uridine diphosphate:UDP)-MurNAc(1)に対してL-Ala,D-Glu, meso-ジアミノピメリン酸(diaminopimelate:DAP),D-Ala-D-Alaが四つのアミノ酸リガーゼ(それぞれMurC, MurD, MurE, MurF)で順次アミド結合することでUDP-MurNAcペンタペプチド(2)が形成される(図1)6, 7).この際必要なD-GluはPLP非依存型のグルタミン酸ラセマーゼで,D-AlaはPLP依存酵素であるアラニンラセマーゼで生合成される8, 9).
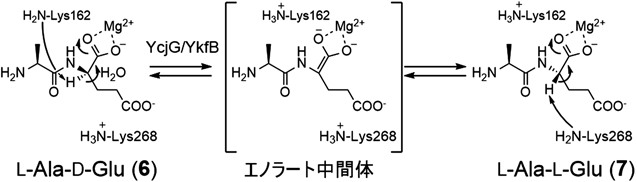
しかし,我々が植物病原菌のザントモナス(Xanthomonas)属細菌のゲノムを精査したところ,UDP-MurNAcペンタペプチド生合成遺伝子のうち,既知のグルタミン酸ラセマーゼ遺伝子のみが見いだせなかった.この事実はXanthomonas属細菌において新規なD-Glu供給系が存在することを示唆している.そこで,グルタミン酸ラセマーゼ遺伝子の欠損に起因するD-Glu要求性大腸菌株のWM335を用いて,D-Glu要求性を相補する遺伝子領域をXanthomonas oryzaeゲノムよりショットガンクローニングで取得したところ,既知酵素と相同性がまったくないXOO_1319,およびMurDと相同性を示すXOO_1320の二つの遺伝子が相補に必要であることが明らかになった.また,組換え酵素を用いたin vitro実験で,XOO_1320がUDP-MurNAc-L-Ala(3)にL-Gluを結合する酵素であり,XOO_1319がUDP-MurNAc-L-Ala-L-Glu(4)の末端のL-Glu残基をエピメリ化しUDP-MurNAc-L-Ala-D-Glu(5)を生成するペプチドのエピメラーゼであることを明らかとした(図1)10).XOO_1320については,大腸菌のMurDと26%相同性があるが,UDP-MurNAc-L-Alaに対して立体化学が異なるL-Gluを付加することからMurD2と命名した.また,MurD2のGlu立体異性体の認識にはArg358とGlu197が重要であり,これら二つのアミノ酸残基を大腸菌由来MurDのものに置換(Arg358Lys, Glu197Asp)するだけで基質特異性をL-GluからD-Gluへ完全に変換できることがわかった11).
XOO_1319は既知酵素と相同性がなく,特徴的なモチーフ配列や補酵素結合領域も持たないまったく新規なエピメラーゼであった.本酵素は,エピメリ化反応に1当量のアデノシン三リン酸(adenosine triphosphate:ATP)とMg2+イオンを必要とし,アデノシン一リン酸(adenosine monophosphate:AMP)を副成する.また,Glu残基のエピメリ化反応には方向性があり,UDP-MurNAc-L-Ala-D-Glu(5)とATPを基質にした反応は進行しなかった.エピメリ化反応では,一般に基質と生成物の自由エネルギーが同程度であるため,両異性体の平衡混合物を与えるが,本酵素はATPの加水分解に伴う自由エネルギーを利用して単方向(L→D)のみのエピメリ化を達成していると考えられる.本酵素はATPを立体反転反応に利用する初の例でありMurLと命名した.反応機構の詳細については今後の解析が待たれるが,基質であるUDP-MurNAc-L-Ala-L-Glu(4)のL-Glu残基のα-カルボン酸をアデニル化により活性化した後にαプロトンの引抜きが起こると考えられる.
ゲノムデータベースを探索したところ,新規ペプチドグリカン生合成経路は,α-プロテオバクテリアであるDevosia属やParvularcula属,γ-プロテオバクテリアであるXylella属,Stenotrophomonas属,Frateuria属,Rhodanobacter属,Pseudoxanthomonas属,Lysobacter属細菌,希少放線菌であるMicromonospora属,Actinoplanes属,Salinispora属細菌などに見いだされ,いずれもMurD2とMurLの相同遺伝子が隣接して存在していた.
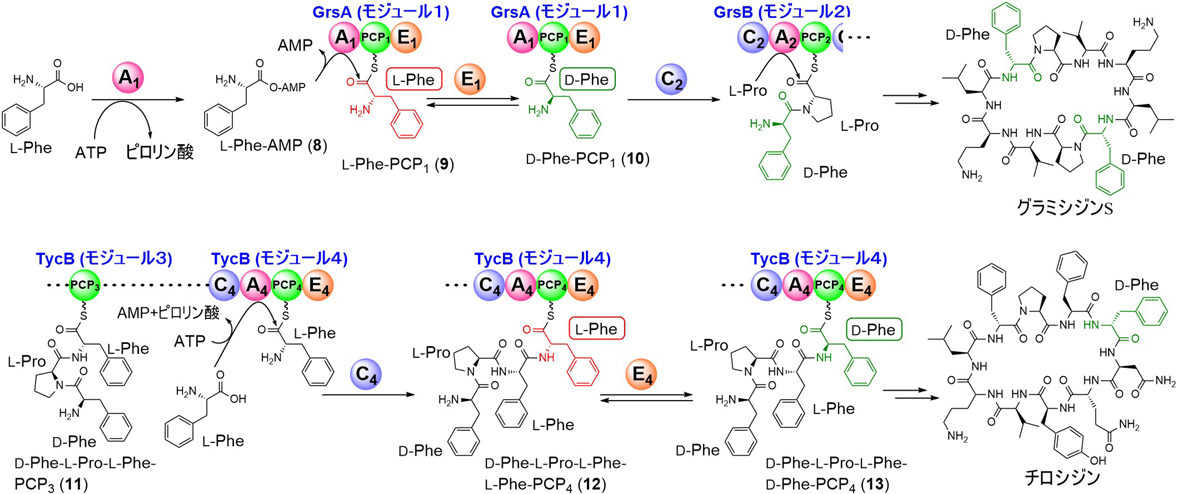
ペニシリンやバンコマイシン,サイクロスポリン等のペプチド系の二次代謝産物の生合成に関わる非リボソームペプチド合成酵素(nonribosomal peptide synthetase:NRPS)は,リボソームによる翻訳機構とはまったく異なるペプチド合成酵素であり,モジュール型の巨大な酵素複合体により逐次的にペプチド鎖を伸長する16).一つのアミノ酸残基の伸長に対応する繰り返し構成単位をモジュールと呼び,その中にはアデニル化(A)ドメイン,縮合(C)ドメイン,ペプチジルキャリアタンパク質(peptidyl carrier protein:PCP)ドメインなどの複数の触媒ドメインが存在する.このうち(A)ドメインは,伸長基質となるアミノ酸の決定因子であり,ATPにより基質アミノ酸のカルボン酸部をアシルアデニル酸として活性化後,PCP上にチオエステル結合を介してロードする.(A)ドメインには,非タンパク質性アミノ酸を用いるものも多く,リボソームのような構成アミノ酸の制限はない.(C)ドメインは,上流のペプチジル-PCPと下流のアミノアシル-PCPの間のアミド結合形成を触媒し,ペプチド鎖を伸長する.上述の基本ドメインに加えて,N-メチル化(NMe)ドメイン,エピメラーゼ(E)ドメイン,チアゾリン/オキサゾリン環形成(Cy)ドメインなどがモジュール中に存在した場合,これらドメインによりペプチド鎖が修飾される.
(E)ドメインは,伸長過程でD-アミノ酸の導入を触媒するが,その触媒機構についてはハーバード大学のWalshらによる実験を皮切りに詳細に解析された17–19).グラミシジンS生合成に関わるNRPSの最初のアミノ酸の取り込みを触媒する開始モジュールには,(A1-PCP1-E1)ドメインが存在し,L-Pheを基質に(PCP1)ドメイン上にD-Pheがロードされる.この過程ではD体の形成反応とアデニル化を経由する(PCP1)ドメインとのアシル化反応が起こるが,組換え酵素とL-Phe, ATPを用いたin vitro解析で反応の順番が詳細に追跡され,反応初期ではL-Pheの消費とともにアデニル体L-Phe-AMP(8)が生成し,続いてL-Phe-PCP1(9)とD-Phe-PCP1(10)が順次生成することが示された17–19).すなわち,(E1)ドメインによるエピメリ化ではL-Phe-PCPチオエステルを基質とする.また,本反応ではd/l-Phe-PCP1の混合物が得られるが,すぐ下流の(C2)ドメインによる縮合でD-Phe-PCP1(10)が選択的にアミド化されることが示されており,最終的には,N末端アミノ酸がd体となったペプチドのみが生合成される(図3)20).また,チロシジンのNRPSの4番目のモジュール(C4-A4-PCP4-E4)では,(A4)ドメインによってL-Pheが(PCP4)ドメインにロードされた後に,(C4)ドメインが上流からの基質であるD-Phe-L-Pro-L-Phe-PCP3(11)とのアミド結合形成反応を触媒してD-Phe-L-Pro-L-Phe-L-Phe-PCP4(12)を生成,最後に(E4)ドメインによるエピメリ化反応で13が進むことが明らかになっている.この場合もエピメリ化がアシル-PCPチオエステル上で進行する点では,グラミシジンSの場合と同様である.X線結晶解析では,保存されたGluとHis残基の側鎖が,それぞれチオエステル基質のαプロトンとカルボニル酸素に近接しており,前者は酸塩基触媒として,後者はエノラートアニオンの安定化に関わることが示されている21).
5. リボソーム翻訳系翻訳後修飾ペプチド生合成に関わるエピメリ化反応
二次代謝産物のD-アミノ酸含有ペプチドの多くは,ペプチド鎖伸長段階での逐次的なエピメリ化が可能なNRPSで生合成される.これに対して,リボソーム由来ペプチドの翻訳後修飾で生合成されるリボソーム翻訳系翻訳後修飾ペプチド(ribosomally synthesized and post-translationally modified peptide:RiPP)と呼ばれる化合物群の中にもD-アミノ酸が見いだされている.RiPPは,N末端側のリーダーペプチドとC末端側のコアペプチドからなる前駆体ペプチドとして翻訳される.前駆体ペプチドの生成後,修飾酵素がリーダーペプチドを認識してコアペプチドの修飾を行うが,この際,前駆体ペプチド認識因子と呼ばれるタンパク質が修飾反応に介在することもある.最終的にリーダーペプチドが切断されて成熟したRiPPが生じる22).RiPP経路におけるD-アミノ酸の導入機構は大きく二つに分けられ,ランチビオティックの生合成にみられSer/Thr残基からの脱水反応と続く1,4-付加反応によるものと,エピメリ化反応によるものがある.後者については,ペプチド基質の位置選択的なアミドαプロトンの引抜きが必要であり,現在までに三つのタイプの酵素が知られている.
1)ランチビオティック生合成
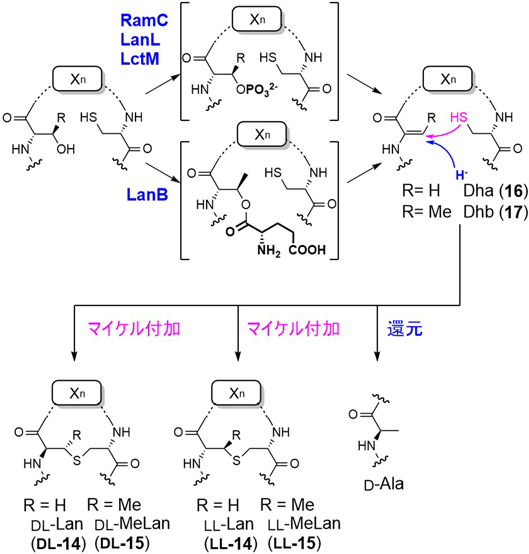
食品保存料として用いられるナイシンに代表されるランチビオティックは,ランチオニン(lanthionine:Lan, 14)やメチルランチオニン(methyllanthionine:MeLan, 15)を含むペプチド抗生物質の総称である23).その生合成については,古くからリボソームで行われることがわかっており,LanはSerとCysに,MeLanはThrとCysに由来することが示されていた.酵素レベルでの解析は,ラクチシン481の生合成が,イリノイ大学のvan der Donkらによって2004年に初めて報告された24).この場合,二つのドメインからなるLctMが,前駆体ペプチドLctAのSer残基の脱水反応(N末端脱水ドメイン)と,生じたデヒドロアラニン(dehydroalanine:Dha, 16)残基に対するCysの硫黄原子のマイケル付加(C末端Zn2+依存環化ドメイン)を触媒してLanが生成する.また,LctMはThrも基質とし,デヒドロブチリン(dehydrobutyrine:Dhb, 17)を経由してMeLanが生成する(図4).なお,脱水反応には1当量のATPが必要であり,Ser/Thrの水酸基がリン酸化された中間体を経由する.また,ナイシンの生合成では,上記とは異なり,Glu-tRNA依存脱水酵素LanBによる水酸基のグルタミル化とβ脱離によりデヒドロアミノ酸が生成する25).この場合,マイケル付加反応は,LctMのC末端側と相同性を示すLanCにより触媒される.なお,Lan形成反応に基づくランチペプチドの分類が提唱されており,LctMタイプはクラスII, LanB/CタイプはクラスIと呼ばれる.また,クラスIIと同様にリン酸化による脱水反応を経由するが,N末端リアーゼドメイン,リン酸化ドメイン,C末端環化ドメインの三つのドメインからなる複合酵素(RamC, LanL)が関与する経路も見いだされ,クラスIIIやIV(環化ドメインによる細分)と呼ばれる26).これらのLan残基の形成反応では,多くの場合DL-Lan(Ser由来のα炭素がD配置で,Cys由来α炭素がL配置)が生成する.この立体選択性は,デヒドロアラニンに対するマイケル付加の段階で決まるが,DL-MeLanとll-MeLanの両者をともに含むシトリシンの生合成酵素を用いた実験で,一つの酵素が両異性体を生成可能なことが見いだされた.また,分子動力学計算や基質類縁体を用いた実験により,Dhb-Dhb-X-X-Cys(Xはデヒドロアミノ酸やCys以外のアミノ酸)配列からはll-MeLanが生成し,それ以外の場合はDL-MeLanとなることが示唆されている27).
また,ランチビオティックの中にはSer残基を由来とするD-Ala残基を含むものが知られている28).D-Alaは,デヒドロアミノ酸に対する1,4-ヒドリド還元により形成されるが,これまで三つのタイプの還元酵素が同定されている(図4).一つ目は,Zn2+依存アルコール脱水素酵素によるもので,シアノバクテリアNostoc punctiformeのNpnJAはNADPHを補酵素にDhaからD-Alaへの還元を触媒することがin vitro実験で示されている29).二つ目は,フラビン依存型の還元酵素で,シトリシン生合成のCrnJやビセレウシン生合成のBsjJBが実験的に同定されている30, 31).また,最近,F420H2依存還元酵素(LxmJ)による還元反応もストレプトミセス属放線菌(Streptomyces rochei)のレクサペプチド生合成で見いだされた32).
2)ラジカルS-アデノシルメチオニン依存ペプチドエピメラーゼ
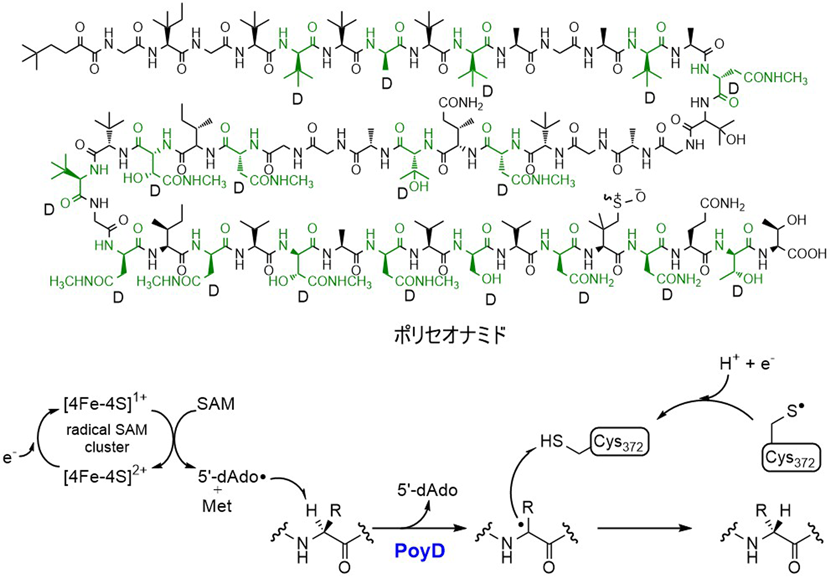
ポリセオナミドは,海綿動物から見いだされたペプチド天然物でtert-ロイシンが8残基,β-ヒドロキシバリンが3残基存在するなど,多くの非タンパク質性アミノ酸を含み,全48アミノ酸残基中18残基がD体である(図5)33).ほとんど非タンパク質性アミノ酸で構成されるため,ポリセオナミドはNRPSによって生合成されると考えられてきた.しかし,スイス連邦工科大学のPielらが,海綿から抽出したメタゲノムから生合成遺伝子を探索した結果,ポリセオナミドのアミノ酸配列に対応する前駆体ペプチド遺伝子(poyA)が見いだされ,驚くべきことに,ポリセオナミドがRiPPであることが示された.なお,後に本遺伝子群は海綿に共生する難培養性のEntotheonella属細菌由来であることが示された34).周辺には9個の翻訳読み枠が存在し,α-ケトグルタル酸依存酸化酵素(poyI)やメチル基転移酵素(poyB, poyC)により,β-ヒドロキシバリンやtert-ロイシンが生成すると考えられた.また,前駆体ペプチド遺伝子に隣接して機能未知のラジカルS-アデノシルメチオニン(S-adenosylmethionine:SAM)酵素遺伝子(poyD)が存在した.ラジカルSAM酵素は,SAMと[4Fe-4S]型の鉄硫黄クラスターを補因子とする酵素であり,[4Fe-4S]+によるSAMの還元的なC–S結合の切断によって生成する5′-デオキシアデノシルラジカル(5′-deoxyadenosyl radical:5′-dAdo・)を利用したラジカル反応を触媒する.本酵素について,poyD遺伝子をpoyA遺伝子と共発現した場合にPoyAにD体アミノ酸残基が導入されたことから,前駆体ペプチドのエピメリ化に関与することが証明された.PoyDは,L→Dの一方向にのみエピメリ化を触媒し,ポリセオナミド構造中のすべてのD-アミノ酸を一つの酵素で導入する35, 36).また,エピメリ化反応の基質認識にはリーダーペプチドが重要であり,基質のC末端からN末端に向けて順次進行することも示されている.PoyDの反応機構については,SAMから生成した5′-dAdo・がエピメリ化するアミノ酸残基のCα水素原子を引き抜き,その後に生じたラジカルが反対側に存在する372番目のCys残基からの水素原子を受け取ることで立体反転が起こると考えられている(図5).本反応は,5′-dAdo・による立体選択的な水素原子の引抜きを利用して,L→Dへの一方向エピメリ化を達成しているが,この方向性により生成物を単一の立体異性体(18残基がD体)へと収束させる例として興味深い.
3)α/β加水分解酵素によるエピメリ化
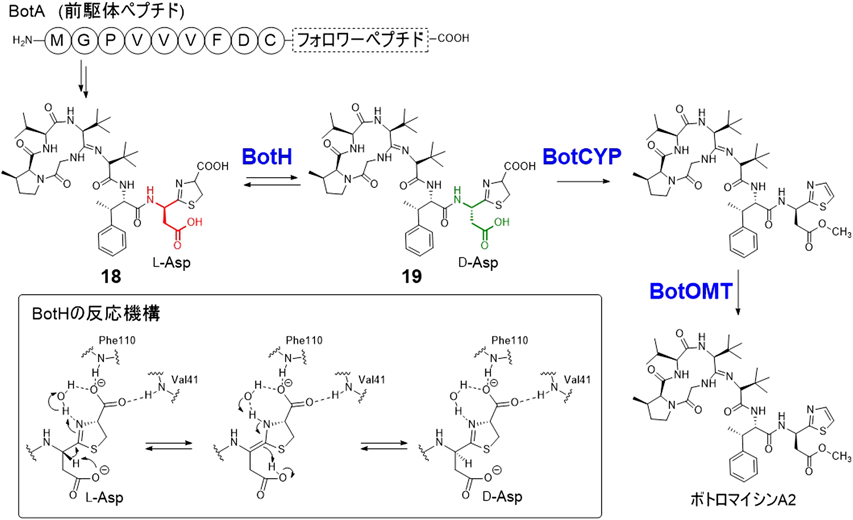
ボトロマイシン類は,Streptomyces bottropensisなどいくつかのストレプトミセス属放線菌から単離され,メチシリン耐性黄色ブドウ球菌を含むグラム陽性細菌に抗菌活性を示す.本化合物群はアミジン構造を介した環状ペプチド構造を特徴とするが,ボトロマイシンA2を含む多くの類縁体はチアゾール環に隣接したD-Ala残基を有する.RiPP経路で生合成されることが示されており,前駆体ペプチドのC-メチル化とチアゾリン環形成,マクロアミジン化後にコアペプチドが切り出されて中間体18が生じ,さらに,L-Asp残基のエピメリ化,脱炭酸,O-メチル化反応により最終産物であるボトロマイシンA2が生成する.L-Asp残基のエピメリ化については,チアゾリン環のα位で起こる反応であり,非酵素的にも18から19が生じることが知られていた(図6).しかし,最近,グラスゴー大学のKoehnkeらは,この反応がα/β加水分解酵素に高い相同性を持つが,ファミリーに保存されている触媒三残基(Ser/His/Asp)を持たない機能未知酵素BotHにより触媒されることを明らかとした37).BotHは,エピメリ化を触媒する初のα/β加水分解酵素であり,可逆反応により二つのエピマーの熱力学的な平衡混合物(18/19)を与える.X線結晶解析でBotHと基質の共結晶が得られており,基質のC末端カルボン酸アニオンとVal41やPhe110のアミド窒素との水素結合が観測された.また,エピメリ化で引き抜かれるAsp残基のαプロトンについては,Asp側鎖カルボン酸と水素結合を形成していた.BotHがチアゾリンに隣接するAspまたはGluのみエピメリ化可能であることもわかっており,本反応は,基質自身が触媒に必要な補助的役割を果たすことが示唆された.すなわち,基質のAsp側鎖カルボン酸が自身のαプロトンを引き抜くことでエナミン中間体が生成し,その後のプロトンが反対側から導入されることでエピメリ化が起こる.なお,本経路では,続くチトクロムP450(BotCYP)による酸化的脱炭酸反応の段階で,エピメリ化後の19が選択されてD-Aspを有する代謝産物のみが生合成される38).
4)ペプチドC末端エピメラーゼ
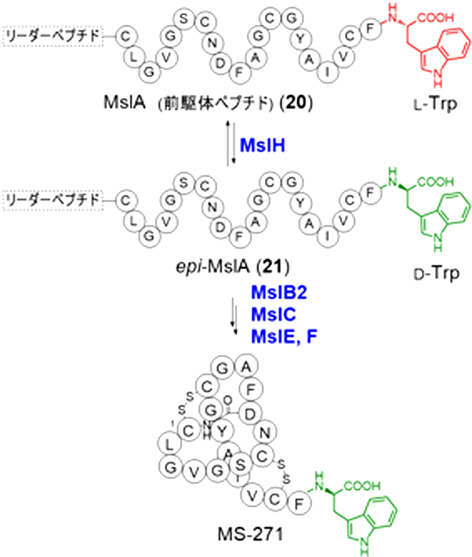
ストレプトミセス属放線菌Streptomyces sp. M-271から単離されたMS-271は21アミノ酸残基からなるラッソペプチド天然物であり,C末端にD-Trp残基を有する(図7).ラッソペプチドは特異な投げ縄構造を持つ天然物の一群で,N末端のアミノ基と8~10残基目に存在するAsp/Gluの側鎖カルボン酸間でのアミド結合を介した環状構造を形成し,C末端側のペプチド鎖がその穴を貫通してロックされた特殊な立体構造をしている39).ラッソペプチドは一般にリボソームで生合成されるため,我々はMS-271のD-Trp残基の導入機構に興味を持ち生合成解析を行った.生産菌のドラフトゲノム解析を行い生合成遺伝子クラスターを同定した結果,MS-271のC末端Trpまでの全配列を含んだ全長42アミノ酸の前駆体ペプチドの遺伝子(mslA)が見いだされたことから,C末端のTrpが翻訳後にエピメリ化されることが示唆された.また,mslA遺伝子の周辺には,投げ縄構造の構築に関わる遺伝子(mslC, mslB1, mslB2)やジスルフィド形成酵素遺伝子(mslE, mslF),機能未知遺伝子(mslH)が存在したものの,既知のラセマーゼやエピメラーゼと相同性を有する遺伝子は見いだせなかった.しかし,近縁放線菌を宿主に用いたクラスターの異種発現実験でMS-271が生産されたことから,我々は,クラスター中の唯一の機能未知酵素であるMslHがTrpのエピメリ化に関与すると推定した40).
生合成におけるエピメリ化の段階は不明であったため,初めに大腸菌を宿主に前駆体ペプチド遺伝子(mslA)をmslHや投げ縄構造構築の遺伝子(mslC, mslB1, mslB2)と共発現し,in vivo解析した.得られたMslAを加水分解後にキラル誘導体化しLC-MSで分析した結果,mslHとの共発現でD-Trpが検出されたことから,MslHが新規エピメラーゼであり,リボソームで生合成された直後の前駆体ペプチド(20)を基質としてepi-MslA(21)を生成することが示された.また,前駆体ペプチド認識因子遺伝子(mslB1)を追加で共発現した場合に21の生成量の増加が確認され,MslHは前駆体ペプチド認識因子を利用する修飾酵素であることが示唆された.また,精製した組換えタンパク質(MslA, MslH, MslB1)を用いたin vitro実験でも,MslHによるMslAからの21の生成と,MslB1による反応促進が確認された.なお,MslHによるエピメリ化は可逆で,長時間の反応でMslA:21が1:1の平衡混合物を生成した.また,リーダーペプチドが基質認識に必須であること,補酵素や金属を必要としないことも示された41).MslHは,補因子非依存的にペプチドC末端アミノ酸残基のエピメリ化を行う初の酵素であり,酸性度が低いカルボン酸のα水素の引抜きが必要であることから,反応機構が興味深い.また,MslHは,コアペプチド部に対する基質認識が緩く,生合成工学を利用した非天然型D-アミノ酸ペプチド創製への利用が期待できる.現在,本酵素の詳細な解明を進めている.