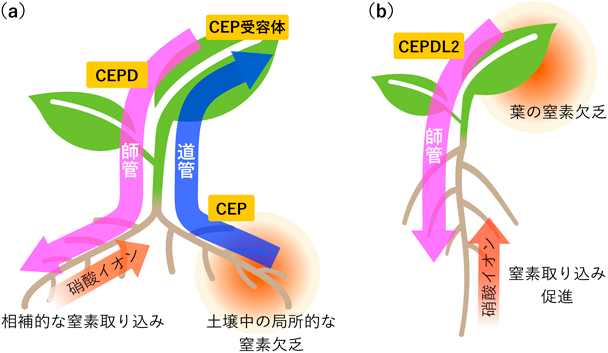
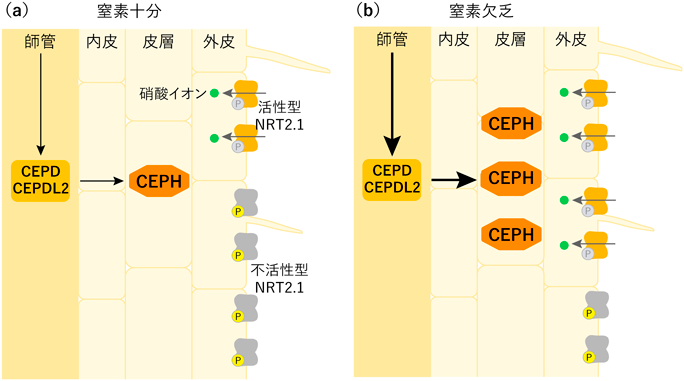
植物の窒素栄養吸収制御の巧みな仕組みAn ingenious mechanism of regulating nitrogen acquisition in plants.
名古屋大学大学院理学研究科生命理学専攻Division of Biological Science, Graduate School of Science, Nagoya University ◇ 〒464–8602 名古屋市千種区不老町 ◇ Furo-cho, Chikusa-ku, Nagoya, Aichi 464–8602, Japan
発行日:2021年6月25日Published: June 25, 2021