2. RIPK1, RIPK3によるTNFシグナルの制御
1)TNFR1複合体(Complex I)形成におけるRIPK1の役割
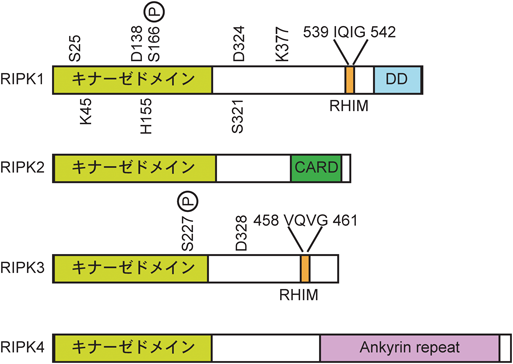
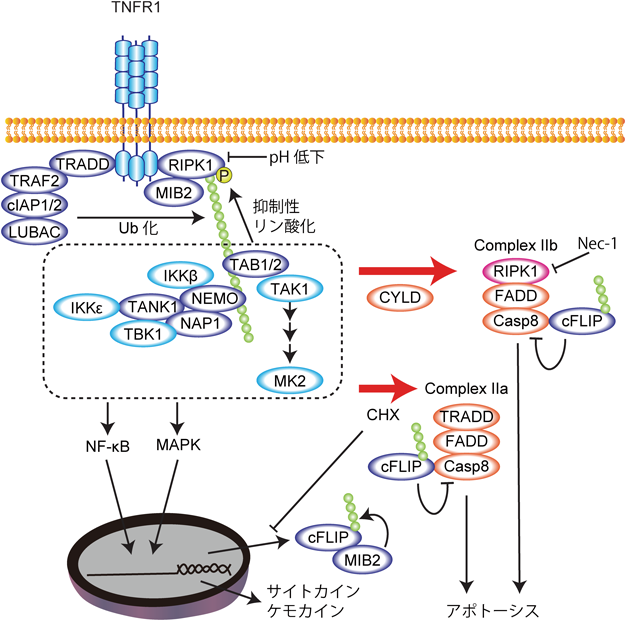
RIPK1は,N末端側にキナーゼドメイン,C末端側にRHIM(RIP homotypic interaction motif)とデスドメインというタンパク質結合ドメインを持つ.この二つのタンパク質結合ドメインを持つがゆえに,RIPK1は非常に多彩な機能を発揮する.以下,その分子機能が最もよく研究されているTNF(tumor necrosis factor)シグナルをモデルとしてRIPK1の機能を概説したい(図2).TNF刺激に応じてRIPK1はデスドメインを介してTNF受容体1(TNFR1)にリクルートされる.TNFR1にはcIAP1(cellular inhibitor of apoptosis 1)やLUBAC(linear ubiquitin chain assembly complex)といったユビキチンリガーゼが同時にリクルートされ,これらの酵素によってRIPK1はK63, M1, K11型などのユビキチン修飾を受ける16–21).このRIPK1に付加されたユビキチン鎖を足場としてIKKβ(inhibitor of nuclear factor-κΒ kinase β),TAK1(transforming growth factor-β-activated kinase 1),IKKε, TBK1(TNFR associated factor family member-associated nuclear factor-κB activator binding kinase 1)といったキナーゼがリクルートされ,Complex Iと呼ばれるTNFR1複合体が形成される22–25).RIPK1のユビキチン化サイトは複数報告されているが26),中でも377番目のリシン(K377)に付加されるK63型ユビキチン鎖はこれらのキナーゼのリクルートに重要であり27),ヒトRIPK1のK377に該当するK376に変異を導入したマウス(Ripk1K376R/K376Rマウス)は過剰なアポトーシスとネクロプトーシスによって胎生13.5日目で致死となる(その理由は後述する)28, 29)(表1).Complex Iにおいて活性化したIKKβとTAK1はそれぞれNF-κΒ(nuclear factor-κB)とMAPK(mitogen-activated protein kinase)経路を活性化し,cFLIP(cellular Fas-associated via death domain-like interleukin 1β converting enzyme inhibitory protein)などの細胞の生存を促す遺伝子やさまざまな炎症性サイトカイン・ケモカインの発現を誘導する.RIPK1を欠損した細胞やシクロヘキシミドなどのタンパク質合成阻害剤で処理をした細胞では,TNFによるこれらの遺伝子発現が抑制され,TNF受容体から解離したTRADD(TNFR superfamily 1A-associated via death domain)がFADD(Fas-associated via death domain),caspase 8(Casp8)と結合して複合体(Complex IIa)を形成し,caspase 8の活性化を引き起こしてアポトーシスを誘導する30–33)(図2).
表1 これまでに報告されているRIPK1変異マウス| 遺伝子 | 遺伝子型 | 影 響 | 生存性 | 表現型のレスキュー | 文 献 |
|---|
| Ripk1 | null | タンパク質欠損 | P0で致死 | Casp8 or FADDとRIPK3 or MLKLの二重欠損によって生存 | 30, 81, 93, 99 |
| TNFR1の単独欠損によって生後1~2週まで生存 |
| Casp8, TRIF, ZBP1の三重欠損によって生存 |
| S25D | IKKβによる抑制性リン酸化の模倣 | 生存(定常状態で異常なし) | | 24 |
| K45A | キナーゼ活性欠損 | 生存(定常状態で異常なし) | | 100 |
| D138N | キナーゼ活性欠損 | 生存(定常状態で異常なし) | | 101 |
| S166A | 自己リン酸化抑制 | 生存(定常状態で異常なし) | | 41 |
| S320D | MK2による抑制性リン酸化の模倣 | 生存(定常状態で異常なし) | | 35 |
| D325A | caspase 8による切断阻害 | E10.5あたりで致死 | Casp8 or FADDとRIPK3 or MLKLの二重欠損によって生存 | 62–65 |
| RIPK1キナーゼ活性欠損(Ripk1D138N)により生後数週まで生存 |
| TNFR1の単独欠損によって生後数週まで生存 |
| K376R | ユビキチン化抑制 | E13.5あたりで致死 | Casp8とRIPK3の二重欠損によって生存 | 28, 29 |
| TNFR1の単独欠損によって生後1~2週まで生存 |
| mRHIM | RHIM変異(539–542 IQIG→AAAA) | P0で致死 | RIPK3, MLKL, ZBP1のいずれかの単独欠損によって生存 | 93, 94 |
2)RIPK1の負の制御機構
RIPK1のリン酸化活性は細胞死誘導能を持つにもかかわらず,多くの細胞において定常状態でのTNF刺激では細胞死は誘導されない.これはなぜであろうか? TNFR1にリクルートされたRIPK1の活性はどのように制御されているのであろうか? この問いに対して多くの研究がなされ,RIPK1のユビキチン鎖にリクルートされた複数のキナーゼがNF-κΒ経路などを活性化するだけでなく,直接RIPK1をリン酸化することで細胞死を抑制していることがわかってきた.Complex Iにおいて,RIPK1のリン酸化活性はIKKβ, TAK1, IKKε, TBK1によってリン酸化されることで抑制されている22–25)(図2).IKKβによる抑制性リン酸化サイトとして25番目のセリン(S25)が同定されており,ここにリン酸化模倣変異(S25D)を導入するとRIPK1のリン酸化活性が顕著に抑制される.一方で,S25A変異を導入してもIKKによるRIPK1リン酸化活性の抑制がみられることから,S25以外にも抑制性リン酸化サイトがあると考えられる.また,Complex IにおいてcIAP1によってRIPK1に付加されたユビキチン鎖自身がRIPK1のリン酸化活性を抑制することも報告されている34).さらに,TAK1の下流で活性化されるMK2(p38/MAPK-activated kinase 2)はComplex Iにはリクルートされないが,細胞質に存在するRIPK1の320番目のセリン(S320)をリン酸化することでRIPK1の活性化を抑制する35–37).
MIB2(mind bomb 2)はRIPK1とともにComplex Iにリクルートされるユビキチンリガーゼであり,そのユビキチン化活性がRIPK1リン酸化活性を阻害し,TNF誘導性細胞死を抑制することが報告されている38).我々もMIB2がRIPK1のリン酸化活性を抑制することを確認しているが,MIB2はComplex I内におけるRIPK1のユビキチン化には関与していなかった.一方で我々は,MIB2がcFLIPと恒常的に結合し,cFLIPをユビキチン化していることを新たに見いだした39).cFLIPはcaspase 8の非活性型ホモログであり,caspase 8に結合してその活性化を抑制するが,MIB2によるcFLIPのユビキチン化がcaspase 8活性化の抑制に重要であった.このことからMIB2は,メカニズムは不明であるがComplex I内でRIPK1のリン酸化活性を阻害し,さらにcFLIPをユビキチン化することでcaspase 8活性化を阻害してTNF誘導性細胞死を抑制すると考えられる.
このようにRIPK1のリン酸化活性はさまざまな分子によって負に制御されている.これらのRIPK1抑制性分子の活性,またはRIPK1のユビキチン化が阻害されると,RIPK1の負の制御が解除され,RIPK1は活性化して自己リン酸化を引き起こす.RIPK1の自己リン酸化サイトは複数同定されているが,特に166番目のセリン(S166)のリン酸化がRIPK1活性化の指標として知られており40),Ripk1S166A/S166AマウスではRIPK1依存的細胞死が抑制される41).活性化したRIPK1はComplex Iから解離し,FADD, caspase 8と結合して複合体(Complex IIb)の形成とcaspase 8の活性化を引き起こしてアポトーシスを誘導する(図2).このComplex IIbによるアポトーシスはRIPK1キナーゼ活性に依存的であり,ネクロスタチンなどのRIPK1キナーゼ活性阻害剤によって抑制される.
RIPK1の活性は上述したリン酸化やユビキチン化以外にもさまざまな機構で制御されている.近年我々は,細胞内pH環境が酸性に傾くとComplex Iの形成には影響を与えないが,RIPK1のキナーゼ活性,Complex IIb形成,そして細胞死が抑制されることを明らかとした.ヒスチジンは生理的条件でみられる程度の酸性化によってプロトン化されて正電荷を帯びる性質があり,pH変化を感知するアミノ酸残基である.RIPK1の155番目のヒスチジン(H155)に変異を導入すると,酸性化によるRIPK1依存的細胞死の抑制がみられなくなったことから,pH変化に伴うRIPK1のプロトン化がRIPK1の活性を抑制する新たな翻訳後修飾である可能性が示された42).H155はATP結合ドメインの近くに位置していることから,正電荷の付加により活性型への構造的遷移が阻害されている可能性が考えられる.細胞内pHの酸性化は低酸素や虚血などの種々の病態で観察されており,RIPK1のpH依存的な活性調節が種々の病態に関与しているかもしれない.
3)RIPK1, RIPK3によるネクロソーム形成の制御
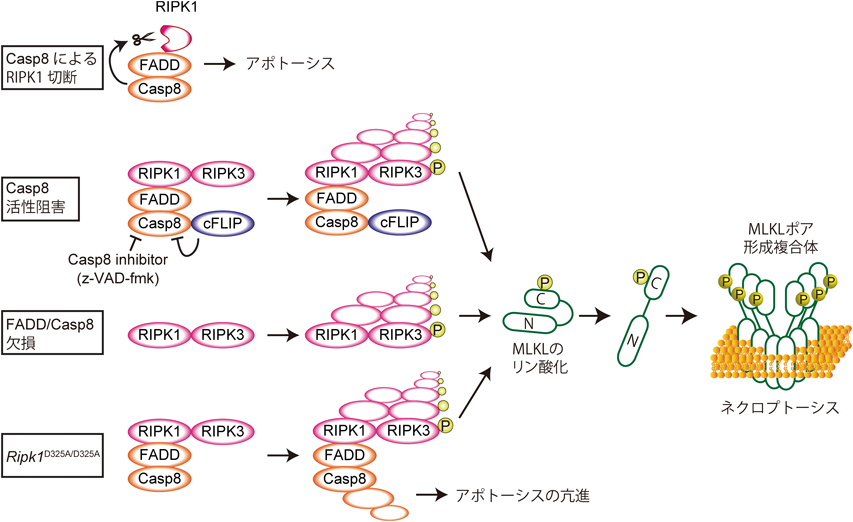
Complex IIa/bが形成される状況でcaspase 8活性を阻害すると,アポトーシスが抑制される代わりに,ネクローシスが引き起こされることが古くに観察されていた43, 44).現在ではこのネクローシスはネクロプトーシスと呼ばれており,その詳細な分子機構が明らかになってきている.
ネクロプトーシスはデス受容体(TNFR1, Fas, TRAIL受容体),Toll様受容体(TLR3, TLR4),インターフェロン(IFN)受容体,T細胞受容体(TCR),核酸受容体(ZBP1)などの刺激によって,RIPK3とその下流分子であるMLKL(mixed lineage kinase domain-like pseudokinase)に依存して引き起こされるネクローシスである45).RIPK3はRIPK1に似てN末端側にキナーゼドメイン,C末端側にRHIMを有するが,デスドメインは有していない(図1).Complex IIa/bが形成される状況でcaspase 8活性を阻害すると,RIPK1とRIPK3は互いのRHIMを介してネクロソームと呼ばれるアミロイド高次複合体を形成する46–53)(図3).このアミロイド複合体におけるRIPK1–RIPK3のストイキオメトリーはいまだ明らかではないが,RIPK1–RIPK3ヘテロアミロイドが形成された後に,RIPK3–RIPK3ホモアミロイドが形成されることで成熟化すると考えられる49).RIPK3はネクロソームの中で自己リン酸化により活性化し,RIPK3の227番目のセリン(S227)が自己リン酸化されると,MLKLと結合してMLKLの357, 358番目のトレオニン,セリン(T357/S358)をリン酸化する54).リン酸化されたMLKLは多量体化して細胞膜を含めた生体膜にポアを形成し,ネクロプトーシスを実行する55–58)(図3).
caspase 8はRIPK159),RIPK360),RIPK1の脱ユビキチン化酵素であるCYLD(cylindromatosis)61)を切断することが知られているが,中でも特にRIPK1の切断がcaspase 8によるネクロプトーシスの抑制に重要であることが最近示された.RIPK1は324番目のアスパラギン酸(D324)でcaspase 8によって切断され,このアスパラギン酸をアラニンに置換したマウス(Ripk1D325A/D325A)では過剰な細胞死が誘導され胎生致死となる(表1).興味深いことに,このマウスはRIPK3を欠損させるだけでは生存せず,RIPK3とcaspase 8を両方欠損させることで生存するようになる.このことから,RIPK1がcaspase 8によって切断されないとComplex IIとネクロソームがより成熟し,活性化したcaspase 8とRIPK3が増加することでアポトーシスとネクロプトーシスの両方が過剰に起こると考えられる62–65).
3. Fas, TRAIL受容体の下流でのRIPK1とRIPK3の働き
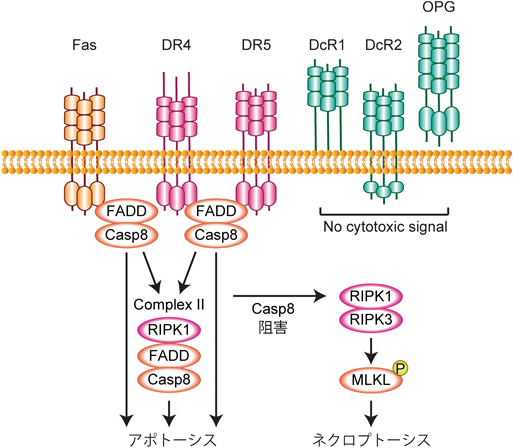
FasとTRAIL受容体(TRAILR)はTNFR1と同じくデス受容体ファミリーに属する.5種類あるTRAIL受容体のうち,デスドメインを持つTRAILR1(DR4)とTRAILR2(DR5)がアポトーシス誘導能を有するのに対して,TRAILR3(DcR1),TRAILR4(DcR2),Osteoprotegerin(OPG)はデスドメインがなくアポトーシスを誘導しない(図4).TNFR1とは異なり,FasとTRAIL受容体のデスドメインにはリガンド依存的にFADDを介してcaspase 8がリクルートされ,細胞膜においてDISC(death-inducing signaling complex)を形成する66).TNFR1 Complex Iと同じく,DISCにはRIPK1, cIAP1, LUBACなどがリクルートされ,NF-κB経路の活性化などが引き起こされるが67),多くの細胞においてFasやTRAIL受容体は第一に細胞死を引き起こす.リガンド刺激から一定時間後にはDISC構成分子は受容体から解離し,細胞質内でComplex IIを形成する.このComplex IIではcaspase 8のさらなる活性化またNF-κB経路の活性化などが引き続き誘導される.FasとTRAIL受容体の下流でも,caspase 8が阻害された状況においては,RIPK1とRIPK3がネクロソームを形成してネクロプトーシスが引き起こされる.
TNFR1におけるComplex IからComplex IIへの移行ならびにFas, TRAIL受容体におけるDISCからComplex IIへの移行を制御する分子機構にはいまだ不明な点が多く残されており,デス受容体シグナルの最大の謎の一つとされている.多くの研究者が細胞内シグナル伝達分子やその翻訳後修飾を解析することでその謎の解明に取り組んでいるが,我々は細胞表面に存在する糖鎖がこの移行を制御する一つの重要な要素であることを明らかとしてきた.フコースは糖鎖を形成する単糖の1種であり,フコースを含むフコシル化糖鎖は発生や分化またがんなどのさまざまな生理的・病理的局面で重要な働きをしている.フコシル化糖鎖の合成にはゴルジ体内での糖転移反応のドナー基質であるGDP-フコースが必須であり,GMDS(GDP-mannose dehydratase)はそのGDP-フコースの合成に必須の酵素である.我々は大腸がん細胞株HCT116やその他のいくつかのがん細胞株,またヒト大腸がん組織でGMDS遺伝子の機能欠損型変異転写産物を見いだした68, 69).さらに,このGMDS変異によりフコシル化糖鎖を欠損しているHCT116細胞に野生型GMDSを導入するとTRAIL受容体やFasを介したアポトーシスが顕著に誘導されるようになり,さらに腫瘍免疫監視機構の一翼を担うNK(natural killer)細胞によって強く殺傷されるようになることを明らかにした69, 70).このとき,TRAIL受容体やFasの直下で形成されるDISC形成には異常がないもののComplex IIの形成とそれに続くアポトーシスが著しく亢進していたことから,細胞表面における糖鎖構造の変化が受容体に質的変化をもたらし,DISC構成成分の受容体からの解離を促進していると考えられる.
4. TRIF, ZBP1を介したネクロプトーシス
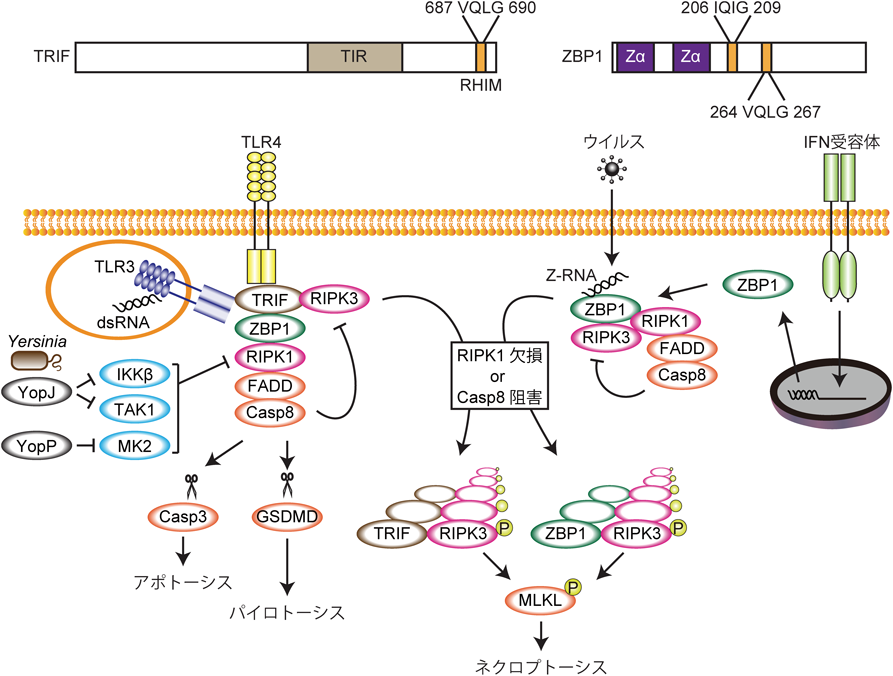
TRIF(Toll/IL-1R domain-containing adaptor inducing interferon β)はTLR3, TLR4の下流でI型IFNの産生を引き起こすアダプター分子である.また,TRIFのC末端側にはRHIMが存在し,このRHIMを介してRIPK1と結合することでNF-κB経路の活性化が引き起こされる71, 72).またTRIF過剰発現によるアポトーシスはTRIF RHIMを介して引き起こされることが報告されている73).最近,マクロファージをTAK1阻害剤の存在下でLPS(lipopolysaccharide)で刺激すると,TRIF-ZBP1-RIPK1-FADD-caspase 8によって形成される複合体(TRIFosome)がRIPK1リン酸化活性依存的に形成され,caspase 8が活性化することが報告された74)(図5).活性化したcaspase 8はcaspase 3/7の切断によるアポトーシスを引き起こすだけでなく,GSDMD(gasdermin D)を切断することでパイロトーシスを引き起こす.また,Yersinia pestisがコードするYopJタンパク質はTAK1とIKKを75, 76),Yersinia enterocoliticaがコードするYopPタンパク質はMK2を阻害する働きを持つため36),これらの細菌に感染した細胞ではRIPK1の負の制御が解除されてRIPK1キナーゼ活性依存的なcaspase 8の活性化とそれによるアポトーシスとパイロトーシスが誘導される77–79).caspase 8の活性化が阻害された状況においては,TRIFはRHIMを介してRIPK3と結合してTRIF-RIPK3ネクロソームを形成し,ネクロプトーシスを引き起こす80)(図5).このように,RIPK1の活性化はアポトーシス,ネクロプトーシスだけでなくパイロトーシス誘導の引き金ともなる.興味深いことにRIPK1を欠損するとTLR3依存的なネクロプトーシスが顕著に亢進することから,RIPK1はTRIF-RIPK3ネクロソームの形成を阻害していると考えられる81, 82).GSDMDはMLKLと似て多量体を形成して細胞膜にポアを形成して細胞の破裂を引き起こす83).最近Ninj1という16 kDaの小さな2回膜貫通型細胞膜タンパク質がGSDMDの下流でパイロトーシスの実行に必要であることが報告された84).Ninj1はネクロプトーシスには不必要であるため,MLKLとGSDMDによるポア形成には違いがあると考えられ,今後のさらなる解析が待たれる.
ZBP1はZ型核酸を認識する細胞質内受容体であり85–88),IFNによってその発現が強く誘導される.ZBP1はN末端側に核酸認識を担う二つのZαドメイン,また中央に二つのRHIMを有している.ZBP1はウイルス感染時に産生されるZ-RNAへの結合に応じてRHIMを介してRIPK3と結合し,ネクロプトーシスを誘導することでウイルス感染に対して防御的に働いている89–92)(図5).また,ZBP1はウイルス感染などがない状況においてもRIPK3と結合してネクロプトーシスを誘導する.RIPK1のRHIMに変異を導入したRipk1RHIM/RHIMマウスがRIPK3とZBP1に依存的にネクロプトーシスを引き起こして生後すぐに死亡することから,RIPK1はRHIM依存的にZBP1-RIPK3ネクロソームによるネクロプトーシスを阻害することが明らかとなった93, 94)(表1).また,表皮特異的Ripk1−/−マウス(Ripk1E-KOマウス)95)また小腸特異的Fadd−/−マウス(FaddIEC-KOマウス)96)はネクロプトーシスを起こして激しい炎症を起こす.Ripk1E-KOマウスだけでなくFaddIEC-KOマウスでの炎症がZBP1 Zαドメイン変異マウスによって改善されたことから,RIPK1が存在していてもcaspase 8活性が阻害されるとZBP1はRIPK3を介してネクロプトーシスを引き起こす97, 98).ZBP1が定常状態において内在性のZ-RNAに結合していることが示されていることから97),RIPK1はRHIM依存的にFAAD-caspase 8をリクルートすることでZBP1-RIPK3ネクロソームの形成とそれによるネクロプトーシスを阻害していると考えられる(図5).以上のように,RIPK1はRIPK3とネクロソームを形成してネクロプトーシスを誘導する一方で,TRIF, ZBP1によるネクロプトーシスを阻害するという働きを持っている.
1)RIPK1による生体恒常性の維持
RIPK1を全身で欠損したマウス(Ripk1−/−マウス)は過剰なアポトーシスとネクロプトーシスによって生後すぐに死亡するが30, 81, 99),RIPK1のキナーゼ活性のみを欠損させたマウス(Ripk1K45A/K45A,Ripk1D138N/D138Nマウス)は正常に生まれ,成長する100, 101)(表1).このことから,上述のとおりRIPK1の足場としての機能がNF-κBなどの生存シグナルの活性化,そしてComplex IIaによるアポトーシスとTRIF/ZBP1-RIPK3ネクロソームによるネクロプトーシスの阻害を通して個体の生存を維持していることがわかる.組織・細胞特異的なRIPK1ノックアウトマウスの解析から,小腸上皮(Villin-Cre, Ripk1IEC-KO)95, 102),皮膚(K14-Cre, Ripk1E-KO)95),造血細胞(Vav-iCre)103),T細胞(Lck-Cre)104),樹状細胞(CD11c-Cre, Ripk1DC-KO)105, 106)などにおいて,RIPK1欠損により過剰な細胞死が引き起こされ,さまざまな障害が引き起こされることがわかっている.最近我々は,Ripk1DC-KOマウスで大腸において樹状細胞の数が減少する一方で好中球と単球の増加が認められること,さらにヒト潰瘍性大腸炎モデルであるデキストラン硫酸(DSS)誘導性腸炎が劇的に抑制されることを見いだした106).これらの表現型はRIPK3を単独で欠損させても変化しないが,FADDとRIPK3を欠損させることで改善したことから,大腸においてはRIPK1が欠損することで樹状細胞にアポトーシスが引き起こされていることが示唆された.同様にRipk1IEC-KOマウスの表現型はRIPK3を単独で欠損させても改善されず,RIPK3とFADDまたはcaspase 8を二重で欠損させることで改善する95, 102).一方で,Ripk1E-KOマウスの表現型はRIPK3単独欠損によって改善される95).これらの結果は,組織・細胞種においてRIPK1欠損によって引き起こされる細胞死の様式が異なることを示している.
Ripk1−/−マウスと異なり,Ripk3−/−マウスは正常に生まれ,発育するため,RIPK3は個体の発生には重要でないと考えられる107).しかし,高齢化したヒトまたはマウスの精巣においてMLKLのリン酸化が観察され,Ripk3−/−オスマウスでは高齢化しても妊孕性が維持されることが報告された108, 109).このことから,ネクロプトーシスが加齢に伴うオスの生殖能力の老化を引き起こしていることが示唆された.ただし,RIPK3の有無にかかわらず高齢化したマウスから産まれた仔の生存率は低かったことから,精巣でネクロプトーシスが起きることが,次世代に異常を及ぼす高齢のオスの妊孕性を低下させ,種の保存に優位性をもたらしているのかもしれない108, 109).
2)RIPK1遺伝子変異とヒト疾患
近年,原発性免疫不全症候群または若年性炎症性腸疾患の患者でRIPK1遺伝子の機能欠失型ホモ変異が同定された110–112).これらの患者は免疫不全による易感染性と腸炎や関節炎といった末梢組織炎症を呈する.RIPK1遺伝子全長にわたって数種類の遺伝子変異が同定されており,どの変異でもRIPK1タンパク質が欠損または劇的に低下している.C末端側のデスドメイン上にある変異でもRIPK1タンパク質の発現が低下しているため,これらはタンパク質安定性に影響を与える変異であると考えられる.興味深いことに,造血幹細胞移植を受けた患者で免疫不全だけでなく,腸炎や関節炎が改善したことが報告されており,ヒトでは免疫細胞に発現するRIPK1が免疫・組織恒常性の維持に重要であることがわかる.一方マウスでは,Ripk1−/−マウスが生後すぐに死亡すること30, 81, 99),Ripk1IEC-KOマウスが激しい腸炎を起こして生後数週間で死亡すること95, 102),Ripk1−/−細胞を骨髄移植したマウスや造血細胞特異的Ripk1−/−マウスでは末梢組織での免疫細胞の浸潤があるものの組織傷害までは認めらないことなどが観察されている99, 103).我々は樹状細胞でRIPK1を欠損させるだけで末梢組織で炎症細胞が増加することを見いだしているが,このマウスにおいても末梢組織傷害が自然発症することはなかった106).このようにヒトとマウスではRIPK1欠損による表現型に違いがある.
RIPK1がD324でcaspase 8によって切断されることを上述したが,近年自己炎症性疾患の患者でRIPK1 D324のヘテロ変異が同定された64, 65).これらの患者は幼少時から周期的な発熱を繰り返し,リンパ節腫脹やその他さまざまな症状を呈する.Ripk1D325/D325マウスは過剰なアポトーシスとネクロプトーシスにより胎生致死となるのに対して(前述),Ripk1D325/+マウスは正常に生まれ,成長する.しかし,TLRリガンドなどの炎症誘導因子を投与すると激しい炎症を引き起こすことから,刺激に対して細胞死を引き起こす感度が非常に高くなっていると考えられる.実際,患者由来末梢血単核球をTNFで刺激するとRIPK1キナーゼ活性依存的にアポトーシスとネクロプトーシスが引き起こされることから,これらの患者でもさまざまな組織・細胞で両方の細胞死が誘導されていると考えられる.
3)感染症におけるRIPK1, RIPK3の役割
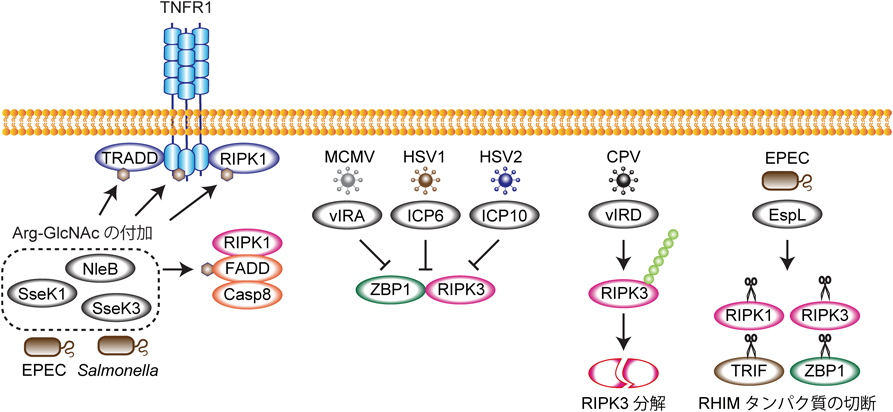
病原体感染において細胞死は,感染細胞の死滅などを通して感染症に対抗する生体防御機構の一翼を担っている.特に,ネクロプトーシス,パイロトーシスといった制御性ネクローシスは感染細胞の死滅と宿主免疫機構の活性化という双方の戦略をもって効率よく病原体を排除することができる.これまでに,RIPK1またはRIPK3がワクシニアウイルス46, 101),ウエストナイルウイルス113),インフルエンザウイルス114),Yersinia菌感染77, 78)などで感染病態に対して防御的な働きをしていることが報告されている.一方で,このような宿主防御機構に抵抗する戦略を有する病原体が見つかってきている(図6).マウスサイトメガロウイルス(mouse cytomegalovirus:MCMV)が有するvIRA(viral inhibitor of RIP activation),そして1型ヘルペスウイルス(herpes simplex virus 1:HSV1),2型ヘルペスウイルス(herpes simplex virus 2:HSV2)がそれぞれ持つICP6, ICP10というタンパク質はRHIMを有しており,ウイルス感染に応答したZBP1-RIPK3ネクロソームによるネクロプトーシスをRHIM依存的に阻害する89, 115, 116).また,牛痘ウイルス(cowpox virus:CPV)やその他のオルソポックスウイルスはvIRD(viral inducer of RIPK3 degradation)というタンパク質を持ち,RIPK3を分解することでネクロプトーシスを阻害している117).微生物感染では,腸管病原性大腸菌(enteropathogenic Escherichia coli: EPEC)が持つタンパク質分解酵素EspLがRHIMを切断することでRHIM含有タンパク質を分解し,RHIM依存的細胞死を抑制している118).また,EPECが持つNleBやサルモネラ菌が持つSseK1, SseK3は,RIPK1をはじめとするデスドメイン含有タンパク質(RIPK1, FADD, TRADD, TNFR1)のデスドメインのアルギニンにN-アセチルグルコサミン(GlcNAc)を付加することでこれらのタンパク質の機能を抑制し,感染細胞の死を阻害している119–124).
このようにRIPK1, RIPK3は病原体感染に対して防御的な働きを持つ一方で,RIPK1リン酸化活性の阻害やRIPK3欠損によって敗血症などでのサイカインストームによる組織傷害が抑制されることが報告されている125, 126).そのため,ひとたび感染が進行した場合はRIPK1, RIPK3は病態を悪化させる原因ともなると考えられる.サイトカインストームは,現在全世界で猛威を奮っているCOVID-19の重症患者でもみられる病態であり,患者の生命を脅かす重大な問題となっている.RIPK1依存的細胞死を引き起こすTNFとIFNγの阻害抗体でCOVID-19の病態が改善することがマウスモデルで示されており127),またCOVID-19患者の肺組織で活性化RIPK1が検出されている128).これらのことからRIPK1の活性抑制がCOVID-19病態の改善につながることが期待され,現在RIPK1阻害剤のCOVID-19に対する臨床試験が進められている.
4)非感染性炎症性疾患におけるRIPK1, RIPK3の役割
これまでに虚血性疾患129–133),神経変性疾患134),皮膚炎100, 135),関節炎136, 137),非アルコール性脂肪肝炎(NASH)138–141)など数多くの非感染性炎症性疾患マウスモデルにおいてRIPK1, RIPK3が病態進展に関与していることが報告されている.これまでに複数の製薬会社が種々のRIPK1リン酸化活性阻害剤を開発し142),炎症性腸疾患,乾癬,関節リウマチ,アルツハイマー病,筋萎縮性側索硬化症に対して臨床試験を行っている.一方で,過去に開発されたRIPK3リン酸化活性阻害剤はネクロプトーシスを阻害するものの,RIPK3のキナーゼドメインを介した二量体化を引き起こしてアポトーシスを誘導することがわかり,治療薬としての開発は進んでいない143, 144).RIPK3はネクロプトーシス経路で特異的に働く分子であると認識されていたが,我々はRIPK3が樹状細胞においてTLR4の下流でNF-κBまたインフラマソームの活性化を制御することでDSS誘導性腸炎における腸組織傷害の修復に寄与していること,またRIPK3-RIPK1-FADD-Caspase 8複合体を形成してアポトーシスを誘導することを明らかにしてきた145–149).現在ではさまざまなグループが,RIPK3がアポトーシス,パイロトーシス,また細胞死以外の経路を制御することを明らかにしており113, 114, 137, 150–152),RIPK3は多様な機能を介して細胞死と炎症を制御していることがわかっている.これらのことから,RIPK1とRIPK3が多くの疾患の病態進展に関わっていることには疑いの余地はないが,両者ともに多様な機能を持つためその病態制御機構については慎重な解析が必要であろう.
RIPK1がネクロプトーシスの制御分子であることが初めて報告されたのが2000年であるが153),それ以来RIPK1阻害剤ネクロスタチン1の開発154),下流分子RIPK346, 47)とMLKL54)の同定と続き,ネクロプトーシスの分子機構が明らかになってきた.また,RIPK1, RIPK3の制御機構の解析を通して,さまざまな受容体の下流で細胞の生死がどのように決定されているかがわかってきた.しかしながら,詳細な分子機構についてはいまだ不明な点が多く残されており,特にRIPK1, RIPK3が関わる分子複合体の形成,活性,解消の制御機構についてはよくわかっていない.この点が明らかになることでRIPK1, RIPK3を標的とした治療薬の開発に新たな道が開けるのではないかと思う.また,RIPK1, RIPK3の解析はほとんどがヒトやマウスを対象にした研究のみであり,進化の過程においてRIPK1, RIPK3がどのような役割を果たしてきたのかは不明である.これは我々がなぜアポトーシス以外にネクロプトーシスやパイロトーシスなどの多様な制御性細胞死を有しているのかという問いかけにもつながるものであり,今後の研究の進展が待たれるところである.
引用文献References
1) Stanger, B.Z., Leder, P., Lee, T.H., Kim, E., & Seed, B. (1995) RIP: A novel protein containing a death domain that interacts with Fas/APO-1 (CD95) in yeast and causes cell death. Cell, 81, 513–523.
2) Inohara, N., del Peso, L., Koseki, T., Chen, S., & Nunez, G. (1998) RICK, a novel protein kinase containing a caspase recruitment domain, interacts with CLARP and regulates CD95-mediated apoptosis. J. Biol. Chem., 273, 12296–12300.
3) Yu, P.W., Huang, B.C., Shen, M., Quast, J., Chan, E., Xu, X., Nolan, G.P., Payan, D.G., & Luo, Y. (1999) Identification of RIP3, a RIP-like kinase that activates apoptosis and NFkappaB. Curr. Biol., 9, 539–542.
4) Holland, P., Willis, C., Kanaly, S., Glaccum, M., Warren, A., Charrier, K., Murison, J., Derry, J., Virca, G., Bird, T., et al. (2002) RIP4 is an ankyrin repeat-containing kinase essential for keratinocyte differentiation. Curr. Biol., 12, 1424–1428.
5) Caruso, R., Warner, N., Inohara, N., & Nunez, G. (2014) NOD1 and NOD2: Signaling, host defense, and inflammatory disease. Immunity, 41, 898–908.
6) Huang, X., McGann, J.C., Liu, B.Y., Hannoush, R.N., Lill, J.R., Pham, V., Newton, K., Kakunda, M., Liu, J., Yu, C., et al. (2013) Phosphorylation of dishevelled by protein kinase RIPK4 regulates Wnt signaling. Science, 339, 1441–1445.
7) Oberbeck, N., Pham, V.C., Webster, J.D., Reja, R., Huang, C.S., Zhang, Y., Roose-Girma, M., Warming, S., Li, Q., Birnberg, A., et al. (2019) The RIPK4-IRF6 signalling axis safeguards epidermal differentiation and barrier function. Nature, 574, 249–253.
8) Xu, J., Wei, Q., & He, Z. (2020) Insight into the function of RIPK4 in keratinocyte differentiation and carcinogenesis. Front. Oncol., 10, 1562.
9) Mitchell, K., O’Sullivan, J., Missero, C., Blair, E., Richardson, R., Anderson, B., Antonini, D., Murray, J.C., Shanske, A.L., Schutte, B.C., et al. (2012) Exome sequence identifies RIPK4 as the Bartsocas-Papas syndrome locus. Am. J. Hum. Genet., 90, 69–75.
10) Kalay, E., Sezgin, O., Chellappa, V., Mutlu, M., Morsy, H., Kayserili, H., Kreiger, E., Cansu, A., Toraman, B., Abdalla, E.M., et al. (2012) Mutations in RIPK4 cause the autosomal-recessive form of popliteal pterygium syndrome. Am. J. Hum. Genet., 90, 76–85.
11) Busa, T., Jeraiby, M., Clemenson, A., Manouvrier, S., Granados, V., Philip, N., & Touraine, R. (2017) Confirmation that RIPK4 mutations cause not only Bartsocas-Papas syndrome but also CHAND syndrome. Am. J. Med. Genet. A., 173, 3114–3117.
12) Zha, J., Zhou, Q., Xu, L.G., Chen, D., Li, L., Zhai, Z., & Shu, H.B. (2004) RIP5 is a RIP-homologous inducer of cell death. Biochem. Biophys. Res. Commun., 319, 298–303.
13) Meylan, E. & Tschopp, J. (2005) The RIP kinases: crucial integrators of cellular stress. Trends Biochem. Sci., 30, 151–159.
14) Taylor, M. & Alessi, D.R. (2020) Advances in elucidating the function of leucine-rich repeat protein kinase-2 in normal cells and Parkinson’s disease. Curr. Opin. Cell Biol., 63, 102–113.
15) 花房 洋,松本邦弘(2017)ROCOファミリーキナーゼLRRK1の基質依存的な細胞機能.生化学,89, 286–289.
16) Bertrand, M.J., Milutinovic, S., Dickson, K.M., Ho, W.C., Boudreault, A., Durkin, J., Gillard, J.W., Jaquith, J.B., Morris, S.J., & Barker, P.A. (2008) cIAP1 and cIAP2 facilitate cancer cell survival by functioning as E3 ligases that promote RIP1 ubiquitination. Mol. Cell, 30, 689–700.
17) Gerlach, B., Cordier, S.M., Schmukle, A.C., Emmerich, C.H., Rieser, E., Haas, T.L., Webb, A.I., Rickard, J.A., Anderton, H., Wong, W.W., et al. (2011) Linear ubiquitination prevents inflammation and regulates immune signalling. Nature, 471, 591–596.
18) Haas, T.L., Emmerich, C.H., Gerlach, B., Schmukle, A.C., Cordier, S.M., Rieser, E., Feltham, R., Vince, J., Warnken, U., Wenger, T., et al. (2009) Recruitment of the linear ubiquitin chain assembly complex stabilizes the TNF-R1 signaling complex and is required for TNF-mediated gene induction. Mol. Cell, 36, 831–844.
19) Ikeda, F., Deribe, Y.L., Skanland, S.S., Stieglitz, B., Grabbe, C., Franz-Wachtel, M., van Wijk, S.J., Goswami, P., Nagy, V., Terzic, J., et al. (2011) SHARPIN forms a linear ubiquitin ligase complex regulating NF-kappaB activity and apoptosis. Nature, 471, 637–641.
20) Silke, J. (2011) The regulation of TNF signalling: What a tangled web we weave. Curr. Opin. Immunol., 23, 620–626.
21) Tokunaga, F., Nakagawa, T., Nakahara, M., Saeki, Y., Taniguchi, M., Sakata, S., Tanaka, K., Nakano, H., & Iwai, K. (2011) SHARPIN is a component of the NF-kappaB-activating linear ubiquitin chain assembly complex. Nature, 471, 633–636.
22) Dondelinger, Y., Jouan-Lanhouet, S., Divert, T., Theatre, E., Bertin, J., Gough, P.J., Giansanti, P., Heck, A.J., Dejardin, E., Vandenabeele, P., et al. (2015) NF-kappaB-independent role of IKKalpha/IKKbeta in preventing RIPK1 Kinase-dependent apoptotic and necroptotic cell death during TNF signaling. Mol. Cell, 60, 63–76.
23) Lafont, E., Draber, P., Rieser, E., Reichert, M., Kupka, S., de Miguel, D., Draberova, H., von Massenhausen, A., Bhamra, A., Henderson, S., et al. (2018) TBK1 and IKKepsilon prevent TNF-induced cell death by RIPK1 phosphorylation. Nat. Cell Biol., 20, 1389–1399.
24) Dondelinger, Y., Delanghe, T., Priem, D., Wynosky-Dolfi, M.A., Sorobetea, D., Rojas-Rivera, D., Giansanti, P., Roelandt, R., Gropengiesser, J., Ruckdeschel, K., et al. (2019) Serine 25 phosphorylation inhibits RIPK1 kinase-dependent cell death in models of infection and inflammation. Nat. Commun., 10, 1729.
25) Xu, D., Jin, T., Zhu, H., Chen, H., Ofengeim, D., Zou, C., Mifflin, L., Pan, L., Amin, P., Li, W., et al. (2018) TBK1 suppresses RIPK1-driven apoptosis and inflammation during development and in aging. Cell, 174, 1477–1491.e19.
26) Simpson, D.S., Gabrielyan, A., & Feltham, R. (2021) RIPK1 ubiquitination: Evidence, correlations and the undefined. Semin. Cell Dev. Biol., 109, 76–85.
27) Ea, C.K., Deng, L., Xia, Z.P., Pineda, G., & Chen, Z.J. (2006) Activation of IKK by TNFalpha requires site-specific ubiquitination of RIP1 and polyubiquitin binding by NEMO. Mol. Cell, 22, 245–257.
28) Zhang, X., Zhang, H., Xu, C., Li, X., Li, M., Wu, X., Pu, W., Zhou, B., Wang, H., Li, D., et al. (2019) Ubiquitination of RIPK1 suppresses programmed cell death by regulating RIPK1 kinase activation during embryogenesis. Nat. Commun., 10, 4158.
29) Kist, M., Komuves, L.G., Goncharov, T., Dugger, D.L., Yu, C., Roose-Girma, M., Newton, K., Webster, J.D., & Vucic, D. (2021) Impaired RIPK1 ubiquitination sensitizes mice to TNF toxicity and inflammatory cell death. Cell Death Differ., 28, 985–1000.
30) Kelliher, M.A., Grimm, S., Ishida, Y., Kuo, F., Stanger, B.Z., & Leder, P. (1998) The death domain kinase RIP mediates the TNF-induced NF-kappaB signal. Immunity, 8, 297–303.
31) Wang, L., Du, F., & Wang, X. (2008) TNF-alpha induces two distinct caspase-8 activation pathways. Cell, 133, 693–703.
32) Anderton, H., Bandala-Sanchez, E., Simpson, D.S., Rickard, J.A., Ng, A.P., Di Rago, L., Hall, C., Vince, J.E., Silke, J., Liccardi, G., et al. (2019) RIPK1 prevents TRADD-driven, but TNFR1 independent, apoptosis during development. Cell Death Differ., 26, 877–889.
33) Dowling, J.P., Alsabbagh, M., Del Casale, C., Liu, Z.G., & Zhang, J. (2019) TRADD regulates perinatal development and adulthood survival in mice lacking RIPK1 and RIPK3. Nat. Commun., 10, 705.
34) Annibaldi, A., Wicky John, S., Vanden Berghe, T., Swatek, K.N., Ruan, J., Liccardi, G., Bianchi, K., Elliott, P.R., Choi, S.M., Van Coillie, S., et al. (2018) Ubiquitin-Mediated Regulation of RIPK1 Kinase Activity Independent of IKK and MK2. Mol. Cell, 69, 566–580.e5.
35) Jaco, I., Annibaldi, A., Lalaoui, N., Wilson, R., Tenev, T., Laurien, L., Kim, C., Jamal, K., Wicky John, S., Liccardi, G., et al. (2017) MK2 Phosphorylates RIPK1 to Prevent TNF-Induced Cell Death. Mol. Cell, 66, 698–710.e5.
36) Menon, M.B., Gropengiesser, J., Fischer, J., Novikova, L., Deuretzbacher, A., Lafera, J., Schimmeck, H., Czymmeck, N., Ronkina, N., Kotlyarov, A., et al. (2017) p38(MAPK)/MK2-dependent phosphorylation controls cytotoxic RIPK1 signalling in inflammation and infection. Nat. Cell Biol., 19, 1248–1259.
37) Dondelinger, Y., Delanghe, T., Rojas-Rivera, D., Priem, D., Delvaeye, T., Bruggeman, I., Van Herreweghe, F., Vandenabeele, P., & Bertrand, M.J.M. (2017) MK2 phosphorylation of RIPK1 regulates TNF-mediated cell death. Nat. Cell Biol., 19, 1237–1247.
38) Feltham, R., Jamal, K., Tenev, T., Liccardi, G., Jaco, I., Domingues, C.M., Morris, O., John, S.W., Annibaldi, A., Widya, M., et al. (2018) Mind bomb regulates cell death during TNF signaling by suppressing RIPK1’s cytotoxic potential. Cell Rep., 23, 470–484.
39) Nakabayashi, O., Takahashi, H., Moriwaki, K., Komazawa-Sakon, S., Ohtake, F., Murai, S., Tsuchiya, Y., Koyahara, Y., Saeki, Y., Yoshida, Y., et al. (2021) MIND bomb 2 prevents RIPK1 kinase activity-dependent and -independent apoptosis through ubiquitylation of cFLIPL. Commun. Biol., 4, 80.
40) Degterev, A., Hitomi, J., Germscheid, M., Ch’en, I.L., Korkina, O., Teng, X., Abbott, D., Cuny, G.D., Yuan, C., Wagner, G., et al. (2008) Identification of RIP1 kinase as a specific cellular target of necrostatins. Nat. Chem. Biol., 4, 313–321.
41) Laurien, L., Nagata, M., Schunke, H., Delanghe, T., Wiederstein, J.L., Kumari, S., Schwarzer, R., Corona, T., Kruger, M., Bertrand, M.J.M., et al. (2020) Autophosphorylation at serine 166 regulates RIP kinase 1-mediated cell death and inflammation. Nat. Commun., 11, 1747.
42) Moriwaki, K., Balaji, S., & Ka-Ming Chan, F. (2020) The death-inducing activity of RIPK1 is regulated by the pH environment. Sci. Signal., 13, eaay7066.
43) Vercammen, D., Beyaert, R., Denecker, G., Goossens, V., Van Loo, G., Declercq, W., Grooten, J., Fiers, W., & Vandenabeele, P. (1998) Inhibition of caspases increases the sensitivity of L929 cells to necrosis mediated by tumor necrosis factor. J. Exp. Med., 187, 1477–1485.
44) Vercammen, D., Brouckaert, G., Denecker, G., Van de Craen, M., Declercq, W., Fiers, W., & Vandenabeele, P. (1998) Dual signaling of the Fas receptor: Initiation of both apoptotic and necrotic cell death pathways. J. Exp. Med., 188, 919–930.
45) Galluzzi, L., Vitale, I., Aaronson, S.A., Abrams, J.M., Adam, D., Agostinis, P., Alnemri, E.S., Altucci, L., Amelio, I., Andrews, D.W., et al. (2018) Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ., 25, 486–541.
46) Cho, Y.S., Challa, S., Moquin, D., Genga, R., Ray, T.D., Guildford, M., & Chan, F.K. (2009) Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell, 137, 1112–1123.
47) He, S., Wang, L., Miao, L., Wang, T., Du, F., Zhao, L., & Wang, X. (2009) Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-alpha. Cell, 137, 1100–1111.
48) Hu, H., Wu, X., Wu, G., Nan, N., Zhang, J., Zhu, X., Zhang, Y., Shu, Z., Liu, J., Liu, X., et al. (2021) RIP3-mediated necroptosis is regulated by inter-filament assembly of RIP homotypic interaction motif. Cell Death Differ., 28, 251–266.
49) Wu, X.L., Hu, H., Dong, X.Q., Zhang, J., Wang, J., Schwieters, C.D., Liu, J., Wu, G.X., Li, B., Lin, J.Y., et al. (2021) The amyloid structure of mouse RIPK3 (receptor interacting protein kinase 3) in cell necroptosis. Nat. Commun., 12, 1627.
50) Mompean, M., Li, W., Li, J., Laage, S., Siemer, A.B., Bozkurt, G., Wu, H., & McDermott, A.E. (2018) The structure of the necrosome RIPK1-RIPK3 core, a human hetero-amyloid signaling complex. Cell, 173, 1244–1253.
51) Wu, X., Ma, Y., Zhao, K., Zhang, J., Sun, Y., Li, Y., Dong, X., Hu, H., Liu, J., Wang, J., et al. (2021) The structure of a minimum amyloid fibril core formed by necroptosis-mediating RHIM of human RIPK3. Proc. Natl. Acad. Sci. USA, 118, e2022933118.
52) Pham, C.L., Shanmugam, N., Strange, M., O’Carroll, A., Brown, J.W., Sierecki, E., Gambin, Y., Steain, M., & Sunde, M. (2019) Viral M45 and necroptosis-associated proteins form heteromeric amyloid assemblies. EMBO Rep., 20, e46518.
53) Li, J., McQuade, T., Siemer, A.B., Napetschnig, J., Moriwaki, K., Hsiao, Y.S., Damko, E., Moquin, D., Walz, T., McDermott, A., et al. (2012) The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis. Cell, 150, 339–350.
54) Sun, L., Wang, H., Wang, Z., He, S., Chen, S., Liao, D., Wang, L., Yan, J., Liu, W., Lei, X., et al. (2012) Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase. Cell, 148, 213–227.
55) Cai, Z., Jitkaew, S., Zhao, J., Chiang, H.C., Choksi, S., Liu, J., Ward, Y., Wu, L.G., & Liu, Z.G. (2014) Plasma membrane translocation of trimerized MLKL protein is required for TNF-induced necroptosis. Nat. Cell Biol., 16, 55–65.
56) Chen, X., Li, W., Ren, J., Huang, D., He, W.T., Song, Y., Yang, C., Zheng, X., Chen, P., & Han, J. (2014) Translocation of mixed lineage kinase domain-like protein to plasma membrane leads to necrotic cell death. Cell Res., 24, 105–121.
57) Dondelinger, Y., Declercq, W., Montessuit, S., Roelandt, R., Goncalves, A., Bruggeman, I., Hulpiau, P., Weber, K., Sehon, C.A., Marquis, R.W., et al. (2014) MLKL compromises plasma membrane integrity by binding to phosphatidylinositol phosphates. Cell Rep., 7, 971–981.
58) Wang, H., Sun, L., Su, L., Rizo, J., Liu, L., Wang, L.F., Wang, F.S., & Wang, X. (2014) Mixed lineage kinase domain-like protein MLKL causes necrotic membrane disruption upon phosphorylation by RIP3. Mol. Cell, 54, 133–146.
59) Lin, Y., Devin, A., Rodriguez, Y., & Liu, Z.G. (1999) Cleavage of the death domain kinase RIP by caspase-8 prompts TNF-induced apoptosis. Genes Dev., 13, 2514–2526.
60) Feng, S., Yang, Y., Mei, Y., Ma, L., Zhu, D.E., Hoti, N., Castanares, M., & Wu, M. (2007) Cleavage of RIP3 inactivates its caspase-independent apoptosis pathway by removal of kinase domain. Cell. Signal., 19, 2056–2067.
61) O’Donnell, M.A., Perez-Jimenez, E., Oberst, A., Ng, A., Massoumi, R., Xavier, R., Green, D.R., & Ting, A.T. (2011) Caspase 8 inhibits programmed necrosis by processing CYLD. Nat. Cell Biol., 13, 1437–1442.
62) Newton, K., Wickliffe, K.E., Dugger, D.L., Maltzman, A., Roose-Girma, M., Dohse, M., Komuves, L., Webster, J.D., & Dixit, V.M. (2019) Cleavage of RIPK1 by caspase-8 is crucial for limiting apoptosis and necroptosis. Nature, 574, 428–431.
63) Zhang, X., Dowling, J.P., & Zhang, J. (2019) RIPK1 can mediate apoptosis in addition to necroptosis during embryonic development. Cell Death Dis., 10, 245.
64) Lalaoui, N., Boyden, S.E., Oda, H., Wood, G.M., Stone, D.L., Chau, D., Liu, L., Stoffels, M., Kratina, T., Lawlor, K.E., et al. (2020) Mutations that prevent caspase cleavage of RIPK1 cause autoinflammatory disease. Nature, 577, 103–108.
65) Tao, P., Sun, J., Wu, Z., Wang, S., Wang, J., Li, W., Pan, H., Bai, R., Zhang, J., Wang, Y., et al. (2020) A dominant autoinflammatory disease caused by non-cleavable variants of RIPK1. Nature, 577, 109–114.
66) Annibaldi, A. & Walczak, H. (2020) Death receptors and their ligands in inflammatory disease and cancer. Cold Spring Harb. Perspect. Biol., 12, a036384.
67) Lafont, E., Kantari-Mimoun, C., Draber, P., De Miguel, D., Hartwig, T., Reichert, M., Kupka, S., Shimizu, Y., Taraborrelli, L., Spit, M., et al. (2017) The linear ubiquitin chain assembly complex regulates TRAIL-induced gene activation and cell death. EMBO J., 36, 1147–1166.
68) Nakayama, K., Moriwaki, K., Imai, T., Shinzaki, S., Kamada, Y., Murata, K., & Miyoshi, E. (2013) Mutation of GDP-mannose-4,6-dehydratase in colorectal cancer metastasis. PLoS One, 8, e70298.
69) Moriwaki, K., Noda, K., Furukawa, Y., Ohshima, K., Uchiyama, A., Nakagawa, T., Taniguchi, N., Daigo, Y., Nakamura, Y., Hayashi, N., et al. (2009) Deficiency of GMDS leads to escape from NK cell-mediated tumor surveillance through modulation of TRAIL signaling. Gastroenterology, 137, 188–198, 198.e1-2.
70) Moriwaki, K., Shinzaki, S., & Miyoshi, E. (2011) GDP-mannose-4,6-dehydratase (GMDS) deficiency renders colon cancer cells resistant to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) receptor- and CD95-mediated apoptosis by inhibiting complex II formation. J. Biol. Chem., 286, 43123–43133.
71) Meylan, E., Burns, K., Hofmann, K., Blancheteau, V., Martinon, F., Kelliher, M., & Tschopp, J. (2004) RIP1 is an essential mediator of Toll-like receptor 3-induced NF-kappa B activation. Nat. Immunol., 5, 503–507.
72) Cusson-Hermance, N., Khurana, S., Lee, T.H., Fitzgerald, K.A., & Kelliher, M.A. (2005) Rip1 mediates the Trif-dependent toll-like receptor 3- and 4-induced NF-{kappa}B activation but does not contribute to interferon regulatory factor 3 activation. J. Biol. Chem., 280, 36560–36566.
73) Kaiser, W.J. & Offermann, M.K. (2005) Apoptosis induced by the toll-like receptor adaptor TRIF is dependent on its receptor interacting protein homotypic interaction motif. J. Immunol., 174, 4942–4952.
74) Muendlein, H.I., Connolly, W.M., Magri, Z., Smirnova, I., Ilyukha, V., Gautam, A., Degterev, A., & Poltorak, A. (2021) ZBP1 promotes LPS-induced cell death and IL-1beta release via RHIM-mediated interactions with RIPK1. Nat. Commun., 12, 86.
75) Mittal, R., Peak-Chew, S.Y., & McMahon, H.T. (2006) Acetylation of MEK2 and I kappa B kinase (IKK) activation loop residues by YopJ inhibits signaling. Proc. Natl. Acad. Sci. USA, 103, 18574–18579.
76) Paquette, N., Conlon, J., Sweet, C., Rus, F., Wilson, L., Pereira, A., Rosadini, C.V., Goutagny, N., Weber, A.N., Lane, W.S., et al. (2012) Serine/threonine acetylation of TGFbeta-activated kinase (TAK1) by Yersinia pestis YopJ inhibits innate immune signaling. Proc. Natl. Acad. Sci. USA, 109, 12710–12715.
77) Weng, D., Marty-Roix, R., Ganesan, S., Proulx, M.K., Vladimer, G.I., Kaiser, W.J., Mocarski, E.S., Pouliot, K., Chan, F.K., Kelliher, M.A., et al. (2014) Caspase-8 and RIP kinases regulate bacteria-induced innate immune responses and cell death. Proc. Natl. Acad. Sci. USA, 111, 7391–7396.
78) Philip, N.H., Dillon, C.P., Snyder, A.G., Fitzgerald, P., Wynosky-Dolfi, M.A., Zwack, E.E., Hu, B., Fitzgerald, L., Mauldin, E.A., Copenhaver, A.M., et al. (2014) Caspase-8 mediates caspase-1 processing and innate immune defense in response to bacterial blockade of NF-kappaB and MAPK signaling. Proc. Natl. Acad. Sci. USA, 111, 7385–7390.
79) Orning, P., Weng, D., Starheim, K., Ratner, D., Best, Z., Lee, B., Brooks, A., Xia, S., Wu, H., Kelliher, M.A., et al. (2018) Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death. Science, 362, 1064–1069.
80) He, S., Liang, Y., Shao, F., & Wang, X. (2011) Toll-like receptors activate programmed necrosis in macrophages through a receptor-interacting kinase-3-mediated pathway. Proc. Natl. Acad. Sci. USA, 108, 20054–20059.
81) Dillon, C.P., Weinlich, R., Rodriguez, D.A., Cripps, J.G., Quarato, G., Gurung, P., Verbist, K.C., Brewer, T.L., Llambi, F., Gong, Y.N., et al. (2014) RIPK1 blocks early postnatal lethality mediated by caspase-8 and RIPK3. Cell, 157, 1189–1202.
82) Kaiser, W.J., Daley-Bauer, L.P., Thapa, R.J., Mandal, P., Berger, S.B., Huang, C., Sundararajan, A., Guo, H., Roback, L., Speck, S.H., et al. (2014) RIP1 suppresses innate immune necrotic as well as apoptotic cell death during mammalian parturition. Proc. Natl. Acad. Sci. USA, 111, 7753–7758.
83) Ruan, J., Xia, S., Liu, X., Lieberman, J., & Wu, H. (2018) Cryo-EM structure of the gasdermin A3 membrane pore. Nature, 557, 62–67.
84) Kayagaki, N., Kornfeld, O.S., Lee, B.L., Stowe, I.B., O’Rourke, K., Li, Q., Sandoval, W., Yan, D., Kang, J., Xu, M., et al. (2021) NINJ1 mediates plasma membrane rupture during lytic cell death. Nature, 591, 131–136.
85) Takaoka, A., Wang, Z., Choi, M.K., Yanai, H., Negishi, H., Ban, T., Lu, Y., Miyagishi, M., Kodama, T., Honda, K., et al. (2007) DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature, 448, 501–505.
86) Ishii, K.J., Kawagoe, T., Koyama, S., Matsui, K., Kumar, H., Kawai, T., Uematsu, S., Takeuchi, O., Takeshita, F., Coban, C., et al. (2008) TANK-binding kinase-1 delineates innate and adaptive immune responses to DNA vaccines. Nature, 451, 725–729.
87) Wang, Z., Choi, M.K., Ban, T., Yanai, H., Negishi, H., Lu, Y., Tamura, T., Takaoka, A., Nishikura, K., & Taniguchi, T. (2008) Regulation of innate immune responses by DAI (DLM-1/ZBP1) and other DNA-sensing molecules. Proc. Natl. Acad. Sci. USA, 105, 5477–5482.
88) Ha, S.C., Kim, D., Hwang, H.Y., Rich, A., Kim, Y.G., & Kim, K.K. (2008) The crystal structure of the second Z-DNA binding domain of human DAI (ZBP1) in complex with Z-DNA reveals an unusual binding mode to Z-DNA. Proc. Natl. Acad. Sci. USA, 105, 20671–20676.
89) Upton, J.W., Kaiser, W.J., & Mocarski, E.S. (2012) DAI/ZBP1/DLM-1 complexes with RIP3 to mediate virus-induced programmed necrosis that is targeted by murine cytomegalovirus vIRA. Cell Host Microbe, 11, 290–297.
90) Maelfait, J., Liverpool, L., Bridgeman, A., Ragan, K.B., Upton, J.W., & Rehwinkel, J. (2017) Sensing of viral and endogenous RNA by ZBP1/DAI induces necroptosis. EMBO J., 36, 2529–2543.
91) Thapa, R.J., Ingram, J.P., Ragan, K.B., Nogusa, S., Boyd, D.F., Benitez, A.A., Sridharan, H., Kosoff, R., Shubina, M., Landsteiner, V.J., et al. (2016) DAI senses influenza a virus genomic RNA and activates RIPK3-dependent cell death. Cell Host Microbe, 20, 674–681.
92) Sridharan, H., Ragan, K.B., Guo, H., Gilley, R.P., Landsteiner, V.J., Kaiser, W.J., & Upton, J.W. (2017) Murine cytomegalovirus IE3-dependent transcription is required for DAI/ZBP1-mediated necroptosis. EMBO Rep., 18, 1429–1441.
93) Newton, K., Wickliffe, K.E., Maltzman, A., Dugger, D.L., Strasser, A., Pham, V.C., Lill, J.R., Roose-Girma, M., Warming, S., Solon, M., et al. (2016) RIPK1 inhibits ZBP1-driven necroptosis during development. Nature, 540, 129–133.
94) Lin, J., Kumari, S., Kim, C., Van, T.M., Wachsmuth, L., Polykratis, A., & Pasparakis, M. (2016) RIPK1 counteracts ZBP1-mediated necroptosis to inhibit inflammation. Nature, 540, 124–128.
95) Dannappel, M., Vlantis, K., Kumari, S., Polykratis, A., Kim, C., Wachsmuth, L., Eftychi, C., Lin, J., Corona, T., Hermance, N., et al. (2014) RIPK1 maintains epithelial homeostasis by inhibiting apoptosis and necroptosis. Nature, 513, 90–94.
96) Welz, P.S., Wullaert, A., Vlantis, K., Kondylis, V., Fernandez-Majada, V., Ermolaeva, M., Kirsch, P., Sterner-Kock, A., van Loo, G., & Pasparakis, M. (2011) FADD prevents RIP3-mediated epithelial cell necrosis and chronic intestinal inflammation. Nature, 477, 330–334.
97) Jiao, H., Wachsmuth, L., Kumari, S., Schwarzer, R., Lin, J., Eren, R.O., Fisher, A., Lane, R., Young, G.R., Kassiotis, G., et al. (2020) Z-nucleic-acid sensing triggers ZBP1-dependent necroptosis and inflammation. Nature, 580, 391–395.
98) Devos, M., Tanghe, G., Gilbert, B., Dierick, E., Verheirstraeten, M., Nemegeer, J., de Reuver, R., Lefebvre, S., De Munck, J., Rehwinkel, J., et al. (2020) Sensing of endogenous nucleic acids by ZBP1 induces keratinocyte necroptosis and skin inflammation. J. Exp. Med., 217, e20191913.
99) Rickard, J.A., O’Donnell, J.A., Evans, J.M., Lalaoui, N., Poh, A.R., Rogers, T., Vince, J.E., Lawlor, K.E., Ninnis, R.L., Anderton, H., et al. (2014) RIPK1 regulates RIPK3-MLKL-driven systemic inflammation and emergency hematopoiesis. Cell, 157, 1175–1188.
100) Berger, S.B., Kasparcova, V., Hoffman, S., Swift, B., Dare, L., Schaeffer, M., Capriotti, C., Cook, M., Finger, J., Hughes-Earle, A., et al. (2014) Cutting Edge: RIP1 kinase activity is dispensable for normal development but is a key regulator of inflammation in SHARPIN-deficient mice. J. Immunol., 192, 5476–5480.
101) Polykratis, A., Hermance, N., Zelic, M., Roderick, J., Kim, C., Van, T.M., Lee, T.H., Chan, F.K.M., Pasparakis, M., & Kelliher, M.A. (2014) Cutting edge: RIPK1 Kinase inactive mice are viable and protected from TNF-induced necroptosis in vivo. J. Immunol., 193, 1539–1543.
102) Takahashi, N., Vereecke, L., Bertrand, M.J., Duprez, L., Berger, S.B., Divert, T., Goncalves, A., Sze, M., Gilbert, B., Kourula, S., et al. (2014) RIPK1 ensures intestinal homeostasis by protecting the epithelium against apoptosis. Nature, 513, 95–99.
103) Roderick, J.E., Hermance, N., Zelic, M., Simmons, M.J., Polykratis, A., Pasparakis, M., & Kelliher, M.A. (2014) Hematopoietic RIPK1 deficiency results in bone marrow failure caused by apoptosis and RIPK3-mediated necroptosis. Proc. Natl. Acad. Sci. USA, 111, 14436–14441.
104) Dowling, J.P., Cai, Y., Bertin, J., Gough, P.J., & Zhang, J. (2016) Kinase-independent function of RIP1, critical for mature T-cell survival and proliferation. Cell Death Dis., 7, e2379.
105) O’Donnell, J.A., Lehman, J., Roderick, J.E., Martinez-Marin, D., Zelic, M., Doran, C., Hermance, N., Lyle, S., Pasparakis, M., Fitzgerald, K.A., et al. (2018) Dendritic cell RIPK1 maintains immune homeostasis by preventing inflammation and autoimmunity. J. Immunol., 200, 737–748.
106) Moriwaki, K., Park, C., Koyama, K., Balaji, S., Kita, K., Yagi, R., Komazawa-Sakon, S., Asuka, T., Nakano, H., Kamada, Y., et al. (2021) The scaffold-dependent function of RIPK1 in dendritic cells promotes injury-induced colitis. bioRxiv.
107) Newton, K., Sun, X., & Dixit, V.M. (2004) Kinase RIP3 is dispensable for normal NF-kappa Bs, signaling by the B-cell and T-cell receptors, tumor necrosis factor receptor 1, and Toll-like receptors 2 and 4. Mol. Cell. Biol., 24, 1464–1469.
108) Li, D., Meng, L., Xu, T., Su, Y., Liu, X., Zhang, Z., & Wang, X. (2017) RIPK1-RIPK3-MLKL-dependent necrosis promotes the aging of mouse male reproductive system. eLife, 6, e27692.
109) Li, D., Ai, Y., Guo, J., Dong, B., Li, L., Cai, G., Chen, S., Xu, D., Wang, F., & Wang, X. (2020) Casein kinase 1G2 suppresses necroptosis-promoted testis aging by inhibiting receptor-interacting kinase 3. eLife, 9, e61564.
110) Cuchet-Lourenco, D., Eletto, D., Wu, C., Plagnol, V., Papapietro, O., Curtis, J., Ceron-Gutierrez, L., Bacon, C.M., Hackett, S., Alsaleem, B., et al. (2018) Biallelic RIPK1 mutations in humans cause severe immunodeficiency, arthritis, and intestinal inflammation. Science, 361, 810–813.
111) Li, Y., Fuhrer, M., Bahrami, E., Socha, P., Klaudel-Dreszler, M., Bouzidi, A., Liu, Y., Lehle, A.S., Magg, T., Hollizeck, S., et al. (2019) Human RIPK1 deficiency causes combined immunodeficiency and inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA, 116, 970–975.
112) Uchiyama, Y., Kim, C.A., Pastorino, A.C., Ceroni, J., Lima, P.P., de Barros Dorna, M., Honjo, R.S., Bertola, D., Hamanaka, K., Fujita, A., et al. (2019) Primary immunodeficiency with chronic enteropathy and developmental delay in a boy arising from a novel homozygous RIPK1 variant. J. Hum. Genet., 64, 955–960.
113) Daniels, B.P., Snyder, A.G., Olsen, T.M., Orozco, S., Oguin, T.H. 3rd, Tait, S.W.G., Martinez, J., Gale, M. Jr., Loo, Y.M., & Oberst, A. (2017) RIPK3 restricts viral pathogenesis via cell death-independent neuroinflammation. Cell, 169, 301–313.e11.
114) Nogusa, S., Thapa, R.J., Dillon, C.P., Liedmann, S., Oguin, T.H. 3rd, Ingram, J.P., Rodriguez, D.A., Kosoff, R., Sharma, S., Sturm, O., et al. (2016) RIPK3 activates parallel pathways of MLKL-Driven necroptosis and FADD-mediated apoptosis to protect against influenza a virus. Cell Host Microbe, 20, 13–24.
115) Guo, H., Omoto, S., Harris, P.A., Finger, J.N., Bertin, J., Gough, P.J., Kaiser, W.J., & Mocarski, E.S. (2015) Herpes simplex virus suppresses necroptosis in human cells. Cell Host Microbe, 17, 243–251.
116) Guo, H., Gilley, R.P., Fisher, A., Lane, R., Landsteiner, V.J., Ragan, K.B., Dovey, C.M., Carette, J.E., Upton, J.W., Mocarski, E.S., et al. (2018) Species-independent contribution of ZBP1/DAI/DLM-1-triggered necroptosis in host defense against HSV1. Cell Death Dis., 9, 816.
117) Liu, Z., Nailwal, H., Rector, J., Rahman, M.M., Sam, R., McFadden, G., & Chan, F.K. (2021) A class of viral inducer of degradation of the necroptosis adaptor RIPK3 regulates virus-induced inflammation. Immunity, 54, 247–258.e7.
118) Pearson, J.S., Giogha, C., Muhlen, S., Nachbur, U., Pham, C.L., Zhang, Y., Hildebrand, J.M., Oates, C.V., Lung, T.W., Ingle, D., et al. (2017) EspL is a bacterial cysteine protease effector that cleaves RHIM proteins to block necroptosis and inflammation. Nat. Microbiol., 2, 16258.
119) Ding, J., Pan, X., Du, L., Yao, Q., Xue, J., Yao, H., Wang, D.C., Li, S., & Shao, F. (2019) Structural and functional insights into host death domains inactivation by the bacterial arginine GlcNAcyltransferase effector. Mol. Cell, 74, 922–935.e6.
120) Gunster, R.A., Matthews, S.A., Holden, D.W., & Thurston, T.L.M. (2017) SseK1 and SseK3 Type III secretion system effectors inhibit NF-kappaB signaling and necroptotic cell death in salmonella-infected macrophages. Infect. Immun., 85, e00010-17.
121) Li, S., Zhang, L., Yao, Q., Li, L., Dong, N., Rong, J., Gao, W., Ding, X., Sun, L., Chen, X., et al. (2013) Pathogen blocks host death receptor signalling by arginine GlcNAcylation of death domains. Nature, 501, 242–246.
122) Park, J.B., Kim, Y.H., Yoo, Y., Kim, J., Jun, S.H., Cho, J.W., El Qaidi, S., Walpole, S., Monaco, S., Garcia-Garcia, A.A., et al. (2018) Structural basis for arginine glycosylation of host substrates by bacterial effector proteins. Nat. Commun., 9, 4283.
123) Pearson, J.S., Giogha, C., Ong, S.Y., Kennedy, C.L., Kelly, M., Robinson, K.S., Lung, T.W., Mansell, A., Riedmaier, P., Oates, C.V., et al. (2013) A type III effector antagonizes death receptor signalling during bacterial gut infection. Nature, 501, 247–251.
124) Xue, J., Hu, S., Huang, Y., Zhang, Q., Yi, X., Pan, X., & Li, S. (2020) Arg-GlcNAcylation on TRADD by NleB and SseK1 is crucial for bacterial pathogenesis. Front. Cell Dev. Biol., 8, 641.
125) Zelic, M., Roderick, J.E., O’Donnell, J.A., Lehman, J., Lim, S.E., Janardhan, H.P., Trivedi, C.M., Pasparakis, M., & Kelliher, M.A. (2018) RIP kinase 1-dependent endothelial necroptosis underlies systemic inflammatory response syndrome. J. Clin. Invest., 128, 2064–2075.
126) Duprez, L., Takahashi, N., Van Hauwermeiren, F., Vandendriessche, B., Goossens, V., Vanden Berghe, T., Declercq, W., Libert, C., Cauwels, A., & Vandenabeele, P. (2011) RIP kinase-dependent necrosis drives lethal systemic inflammatory response syndrome. Immunity, 35, 908–918.
127) Karki, R., Sharma, B.R., Tuladhar, S., Williams, E.P., Zalduondo, L., Samir, P., Zheng, M., Sundaram, B., Banoth, B., Malireddi, R.K.S., et al. (2021) Synergism of TNF-alpha and IFN-gamma triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndromes. Cell, 184, 149–168.e17.
128) Riebeling, T., Jamal, K., Wilson, R., Kolbrink, B., von Samson-Himmelstjerna, F.A., Moerke, C., Ramos Garcia, L., Dahlke, E., Michels, F., Luhder, F., et al. (2021) Primidone blocks RIPK1-driven cell death and inflammation. Cell Death Differ., 28, 1610–1626.
129) Zhang, Y., Li, M., Li, X., Zhang, H., Wang, L., Wu, X., Zhang, H., & Luo, Y. (2020) Catalytically inactive RIP1 and RIP3 deficiency protect against acute ischemic stroke by inhibiting necroptosis and neuroinflammation. Cell Death Dis., 11, 565.
130) Naito, M.G., Xu, D., Amin, P., Lee, J., Wang, H., Li, W., Kelliher, M., Pasparakis, M., & Yuan, J. (2020) Sequential activation of necroptosis and apoptosis cooperates to mediate vascular and neural pathology in stroke. Proc. Natl. Acad. Sci. USA, 117, 4959–4970.
131) Linkermann, A., Brasen, J.H., Himmerkus, N., Liu, S., Huber, T.B., Kunzendorf, U., & Krautwald, S. (2012) Rip1 (receptor-interacting protein kinase 1) mediates necroptosis and contributes to renal ischemia/reperfusion injury. Kidney Int., 81, 751–761.
132) Linkermann, A., Brasen, J.H., Darding, M., Jin, M.K., Sanz, A.B., Heller, J.O., De Zen, F., Weinlich, R., Ortiz, A., Walczak, H., et al. (2013) Two independent pathways of regulated necrosis mediate ischemia-reperfusion injury. Proc. Natl. Acad. Sci. USA, 110, 12024–12029.
133) Stark, K., Goncharov, T., Varfolomeev, E., Xie, L., Ngu, H., Peng, I., Anderson, K.R., Verschueren, E., Choi, M., Kirkpatrick, D.S., et al. (2021) Genetic inactivation of RIP1 kinase activity in rats protects against ischemic brain injury. Cell Death Dis., 12, 379.
134) Yuan, J., Amin, P., & Ofengeim, D. (2019) Necroptosis and RIPK1-mediated neuroinflammation in CNS diseases. Nat. Rev. Neurosci., 20, 19–33.
135) Duan, X., Liu, X., Liu, N., Huang, Y., Jin, Z., Zhang, S., Ming, Z., & Chen, H. (2020) Inhibition of keratinocyte necroptosis mediated by RIPK1/RIPK3/MLKL provides a protective effect against psoriatic inflammation. Cell Death Dis., 11, 134.
136) Jhun, J., Lee, S.H., Kim, S.Y., Ryu, J., Kwon, J.Y., Na, H.S., Jung, K., Moon, S.J., Cho, M.L., & Min, J.K. (2019) RIPK1 inhibition attenuates experimental autoimmune arthritis via suppression of osteoclastogenesis. J. Transl. Med., 17, 84.
137) Lawlor, K.E., Khan, N., Mildenhall, A., Gerlic, M., Croker, B.A., D’Cruz, A.A., Hall, C., Kaur Spall, S., Anderton, H., Masters, S.L., et al. (2015) RIPK3 promotes cell death and NLRP3 inflammasome activation in the absence of MLKL. Nat. Commun., 6, 6282.
138) Tao, L., Yi, Y., Chen, Y., Zhang, H., Orning, P., Lien, E., Jie, J., Zhang, W., Xu, Q., Li, Y., et al. (2021) RIP1 kinase activity promotes steatohepatitis through mediating cell death and inflammation in macrophages. Cell Death Differ.28, 1418–1433.
139) Majdi, A., Aoudjehane, L., Ratziu, V., Islam, T., Afonso, M.B., Conti, F., Mestiri, T., Lagouge, M., Foufelle, F., Ballenghien, F., et al. (2020) Inhibition of receptor-interacting protein kinase 1 improves experimental non-alcoholic fatty liver disease. J. Hepatol., 72, 627–635.
140) Gautheron, J., Vucur, M., Reisinger, F., Cardenas, D.V., Roderburg, C., Koppe, C., Kreggenwinkel, K., Schneider, A.T., Bartneck, M., Neumann, U.P., et al. (2014) A positive feedback loop between RIP3 and JNK controls non-alcoholic steatohepatitis. EMBO Mol. Med., 6, 1062–1074.
141) Afonso MB, Rodrigues PM, Mateus-Pinheiro M, Simao AL, Gaspar MM, Majdi A, Arretxe E, Alonso C, Santos-Laso A, Jimenez-Aguero R, et al. (2020) RIPK3 acts as a lipid metabolism regulator contributing to inflammation and carcinogenesis in non-alcoholic fatty liver disease. Gut., gutjnl-2020-321767.
142) Mifflin, L., Ofengeim, D., & Yuan, J. (2020) Receptor-interacting protein kinase 1 (RIPK1) as a therapeutic target. Nat. Rev. Drug Discov., 19, 553–571.
143) Mandal, P., Berger, S.B., Pillay, S., Moriwaki, K., Huang, C., Guo, H., Lich, J.D., Finger, J., Kasparcova, V., Votta, B., et al. (2014) RIP3 induces apoptosis independent of pronecrotic kinase activity. Mol. Cell, 56, 481–495.
144) Raju, S., Whalen, D.M., Mengistu, M., Swanson, C., Quinn, J.G., Taylor, S.S., Webster, J.D., Newton, K., & Shaw, A.S. (2018) Kinase domain dimerization drives RIPK3-dependent necroptosis. Sci. Signal., 11, eaar2188.
145) Moriwaki, K., Balaji, S., McQuade, T., Malhotra, N., Kang, J., & Chan, F.K. (2014) The necroptosis adaptor RIPK3 promotes injury-induced cytokine expression and tissue repair. Immunity, 41, 567–578.
146) Moriwaki, K., Bertin, J., Gough, P.J., & Chan, F.K.M. (2015) A RIPK3-caspase 8 complex mediates atypical pro-IL-1b processing. J. Immunol., 194, 1938–1944.
147) Moriwaki, K. & Chan, F.K. (2016) Regulation of RIPK3 and RHIM-dependent necroptosis by the proteasome. J. Biol. Chem., 291, 5948–5959.
148) Moriwaki, K. & Chan, F.K. (2017) The inflammatory signal adaptor RIPK3: Functions beyond necroptosis. Int. Rev. Cell Mol. Biol., 328, 253–275.
149) Moriwaki, K., Balaji, S., Bertin, J., Gough, P.J., & Chan, F.K. (2017) Distinct kinase-independent role of RIPK3 in CD11c(+) mononuclear phagocytes in cytokine-induced tissue repair. Cell Rep., 18, 2441–2451.
150) Alvarez-Diaz, S., Dillon, C.P., Lalaoui, N., Tanzer, M.C., Rodriguez, D.A., Lin, A., Lebois, M., Hakem, R., Josefsson, E.C., O’Reilly, L.A., et al. (2016) The pseudokinase MLKL and the kinase RIPK3 have distinct roles in autoimmune disease caused by loss of death-receptor-induced apoptosis. Immunity, 45, 513–526.
151) Najjar, M., Saleh, D., Zelic, M., Nogusa, S., Shah, S., Tai, A., Finger, J.N., Polykratis, A., Gough, P.J., Bertin, J., et al. (2016) RIPK1 and RIPK3 kinases promote cell-death-independent inflammation by toll-like receptor 4. Immunity, 45, 46–59.
152) Samir, P., Malireddi, R.K.S., & Kanneganti, T.D. (2020) The PANoptosome: A deadly protein complex driving pyroptosis, apoptosis, and necroptosis (PANoptosis). Front. Cell. Infect. Microbiol., 10, 238.
153) Holler, N., Zaru, R., Micheau, O., Thome, M., Attinger, A., Valitutti, S., Bodmer, J.L., Schneider, P., Seed, B., & Tschopp, J. (2000) Fas triggers an alternative, caspase-8-independent cell death pathway using the kinase RIP as effector molecule. Nat. Immunol., 1, 489–495.
154) Degterev, A., Huang, Z., Boyce, M., Li, Y., Jagtap, P., Mizushima, N., Cuny, G.D., Mitchison, T.J., Moskowitz, M.A., & Yuan, J. (2005) Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury. Nat. Chem. Biol., 1, 112–119.