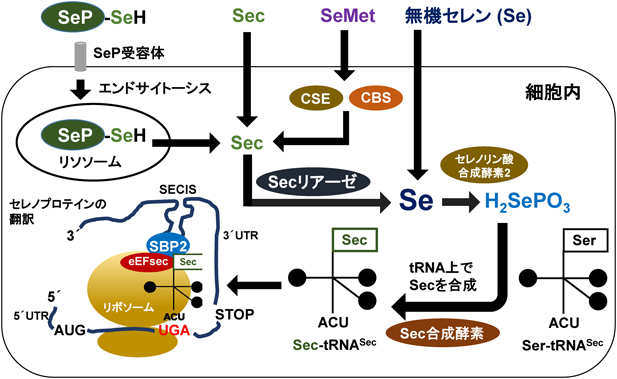
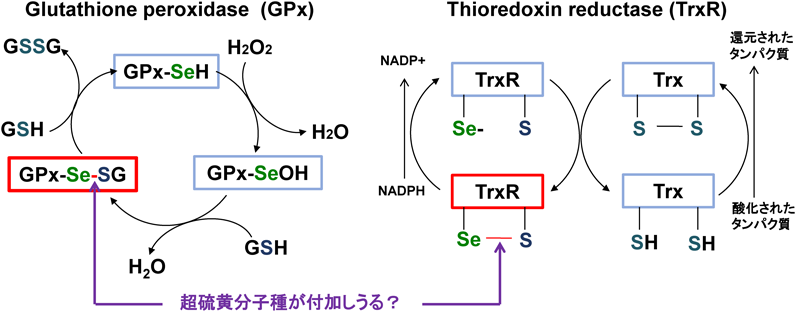
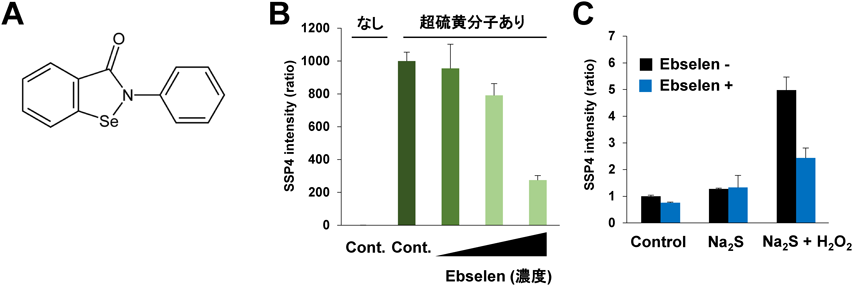
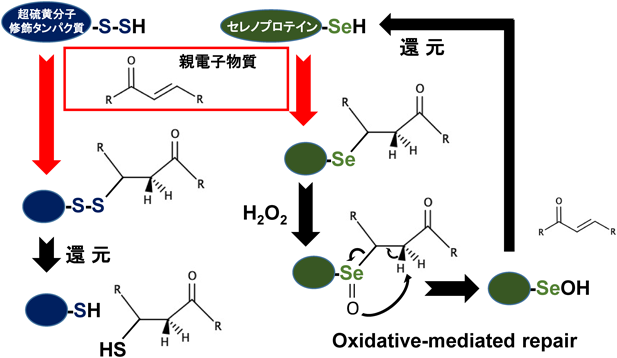
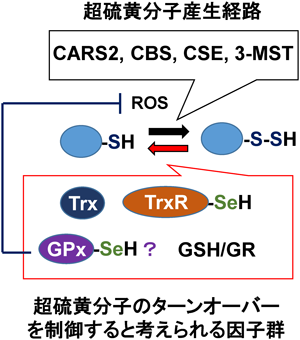
セレンによる超硫黄分子ターンオーバーTurnover of super sulfide species by selenium
東北大学大学院薬学研究科Graduate School of Pharmaceutical Sciences, Tohoku University ◇ 〒980–8578 宮城県仙台市青葉区荒巻字青葉6–3 ◇ 6–3 Aoba, Aramaki, Aoba-ku, Sendai, Miyagi 980–8578, Japan
発行日:2021年10月25日Published: October 25, 2021