ユビキチン分子によるコーディングシステムThe ubiquitin coding system
九州大学生体防御医学研究所Medical Institute of Bioregulation, Kyushu University ◇ 〒812–8582 福岡県福岡市東区馬出3–1–1 ◇ 3–1–1 Maidashi, Higashi-ku, Fukuoka 812–8582, Japan
発行日:2021年12月25日Published: December 25, 2021
ユビキチン分子は,タンパク質の翻訳後修飾に使われる.その名のとおりユビキタスに発現し,ヒトにおいては少なくとも5000の標的基質が存在するとされる.また,ユビキチン化を誘導する酵素は,ヒトでは600以上とされることから,その複雑な制御メカニズムが容易に推測できる.ユビキチンはさまざまなコーディングシステムを持ち,目的によっては,分解系タグ,もしくは非分解系のタグとして機能する.ユビキチン化制御メカニズムの理解は,それらの制御する生物学的効果の理解にもつながることから,構造学的視点から生物学的視点までを広くカバーする研究が展開されている.本稿では,ユビキチンコードがどのように細胞内シグナルを制御するのかについて,炎症制御に着眼して最新の知見と考察を述べる.
© 2021 公益社団法人日本生化学会© 2021 The Japanese Biochemical Society
ユビキチンには,よく知られたプロテアソーム依存性の標的基質分解機能だけでなく,多岐にわたる機能がある.本稿では,ユビキチンによる翻訳後修飾(ユビキチン化)の基本的な概念と,ユビキチン化の炎症性細胞内シグナル,および細菌感染時における役割について着眼する.ユビキチン化の中でも特に,非典型的なユビキチン結合型である直鎖型,さらには最近その機能が明らかにされつつあるエステル結合型についても解説する.
ユビキチンは酵母からヒトまで保存された8.6 kDaの小分子である(図1a).ヒトにおいては,ユビキチンをコードする遺伝子は四つ存在する1, 2).ユビキチン分子は標的基質を翻訳後修飾,つまりユビキチン化することにより,細胞内のあらゆる効果を制御しているといってよい3).リン酸化やアセチル化といった翻訳後修飾とユビキチン化の大きな違いの一つは,その複雑で多様なユビキチン化の「型」(ユビキチンコード)である4).ユビキチンの持つ七つのリシン基,およびN末端のメチオニン基を標的に,ユビキチン分子がユビキチン分子をユビキチン化できる(図1a)5).ユビキチンは,主にリシン基(K)をユビキチン化部位として結合する(典型的結合部位)が,特異的な酵素の働きによっては,N末端のメチオニン基を標的にした非典型的結合部位を介した直鎖型(M1連結型)ユビキチン鎖も形成できる.最近では,ユビキチン分子内のセリン基(S),トレオニン基(T)を標的にしたオキシエステル結合によるユビキチン鎖を形成することも明らかにされた6–9).さらには,ユビキチン分子自身がリン酸化やアセチル化といった翻訳後修飾を受けることにより,ユビキチン化の「型」は複雑性を増す(図1b)5).理論的には,長さの異なる枝分かれや混合結合型を含む,さまざまな型のユビキチンの鎖を基質上に同時に形成することが可能である(図1c).実際に,哺乳類細胞内では,いくつかの結合型のユビキチン鎖が,標的基質の複数のユビキチン化部位において同時に形成されることも観察されている.
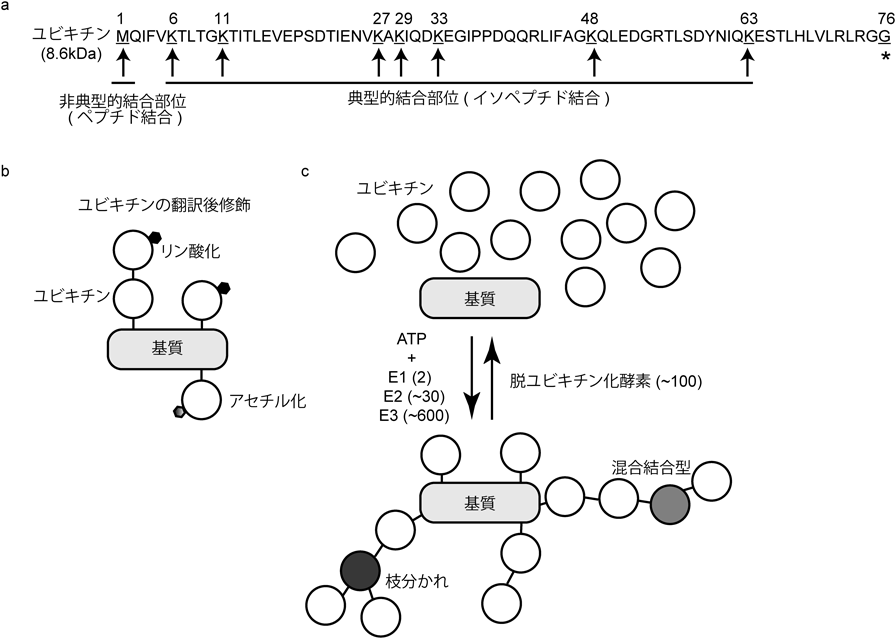
(a)ユビキチン分子(ヒト)のアミノ酸配列,およびユビキチン鎖形成における結合部位.8.6 kDaであるユビキチン分子はC末端のグリシン(G*)が,他のユビキチン分子の結合部位とイソペプチド結合,もしくはペプチド結合することにより,異なる結合型のユビキチン鎖を形成できる.(b)翻訳後修飾されたユビキチンによる基質のユビキチン化.ユビキチン分子自身もリン酸化やアセチル化といった翻訳後修飾を受けることが可能である.(c)基質のユビキチン化と制御する酵素.ユビキチン化においては,まずユビキチンがATP依存性にユビキチン活性化酵素(E1)により活性化される.そして,ヒトにおいて約30種あるとされるユビキチン結合酵素(E2),約600種あるとされるユビキチンリガーゼ(E3)によって,特定の基質がさまざまな結合型のユビキチン鎖によって,ユビキチン化される.また,ユビキチン鎖は100ほど存在するとされる脱ユビキチン化酵素によって,加水分解される.
ユビキチン化は,ATP依存性の3段階の酵素活性により誘導される10).まず,ATPによりユビキチン活性化酵素(E1)がユビキチンを活性化し,そのユビキチンがユビキチン結合酵素(E2)へ転移,そして基質へはユビキチンリガーゼ(E3)が結合させる(図1c).ユビキチンの型や,基質のユビキチン化部位の選択は,約30のE2と約600のE3のペア依存的に行われるとされるが11),その機構には未解明の部分も多く残されている.
ユビキチン化は可逆性の反応であり,ヒトにおいて100ほど存在するとされる脱ユビキチン化酵素が,結合型に対して特異的に,もしくはより柔軟性をもってユビキチン鎖を加水分解する2).ユビキチン化に関わる酵素,および脱ユビキチン化酵素は,アミノ酸配列のバイオインフォマティクス解析により,活性化部位を特定して分類されてきた経緯があることから,実際に酵素活性を持つのかについても不明なものが多い.ただし,最近では,活性化プローブと質量分析を組み合わせた方法から,バイオインフォマティクス解析では同定されてこなかった酵素も発見されており8),今後新たにユビキチン化や脱ユビキチン化を誘導する酵素が同定されることが大いに期待される.
このように,細胞内では基質のさまざまな型のユビキチン化が可能であるが,ユビキチン化による基質への直接的な影響とは何であろうか? 一つには,ユビキチン化により,基質に形成されたタンパク質結合表面が,ユビキチン結合ドメイン(ubiquitin binding domain:UBD)を持つタンパク質を偏在させることがあげられる(図2a)12).ユビキチン化の一番よく知られている機能,つまりプロテアソーム依存性基質分解も13),基質に結合したK48結合型のユビキチン鎖がプロテアソームの蓋を構成する分子と会合することにより誘導される12).これまでに約20のUBDが知られており,ある特定の型のユビキチン鎖を認識するものや,より柔軟な選択肢をもってユビキチン鎖を認識するものなど,さまざまなスタイルがある14).たとえば,ubiquitin binding in ABIN and NEMO(UBAN)というUBDは,NF-κB essential modulator(NEMO)をはじめ,五つの分子に存在する15, 16).これらすべての分子のUBANドメインは,誘導された直鎖型ユビキチン鎖を特異的に認識し,それによって下流の細胞内シグナルを制御する.
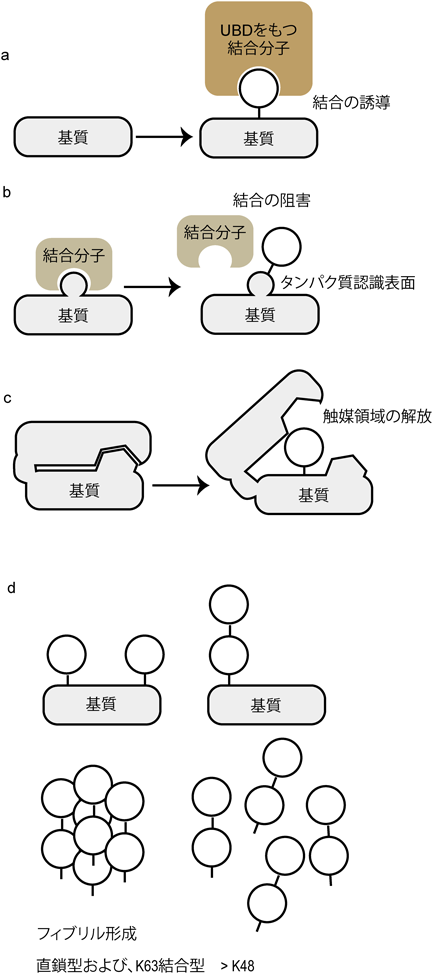
(a, b)基質のユビキチン化により,結合タンパク質のリクルートもしくは阻害が起こる.ユビキチン結合部位(UBD)を持つパートナータンパク質が,ユビキチン依存性にリクルートされる(a).一方,基質が結合部位特異的にユビキチン化されることにより,その結合が阻害されるケースも考えられる(b).(c)基質(不活性型)がユビキチン化されることによって,構造学的変化の誘導に伴い活性化される.(d) 2分子のユビキチンによって修飾を受ける基質においても,そのパターンにより影響は異なる(左:2か所の単量体ユビキチン化,右:1か所の二量体ユビキチン化).可溶性の高い単量体と比較して,二量体の場合は結合型によって可溶性が異なることから,基質分子に対する影響も異なる.直鎖型と,K63結合型のユビキチン二量体は,K48連結型の二量体より可溶性が低く,繊維化する傾向が高い.
二つ目としては,ユビキチン化が,基質の結合分子認識表面に起こることによって,その会合を阻害するケースが考えられる(図2b).さらには,ユビキチン化が基質タンパク質の構造変化を誘導したり,ユビキチンとのアロステリックな結合が構造変化を誘導したりすることによって,不活性型の酵素を活性化することも可能である(図2c)17, 18).これらの三つの場合(図2a~c)は,ユビキチン化が基質のある特定の部位を標的にすることにより制御される.
一方で,ユビキチン化の影響が,より全体的に誘導されるケースも考えられる.たとえば,ユビキチン分子は単量体では可溶性も高く安定しているが,2分子長のユビキチン鎖を形成すると,その生化学的特徴が変化する19).炎症性細胞内シグナル伝達に重要である直鎖型ユビキチン鎖は,細胞内で分解系タグとして機能するK48結合型ユビキチン鎖と比較して,繊維化しやすいこと,またその特徴は鎖の長さに依存して大きくなることが生化学的に示されている(図2d)19).よって,基質に結合するユビキチン鎖の型と,その長さ,同時に起こる結合部位の数により,ユビキチンを介した基質の細胞内の状態が大きく変わると予測される.このような制御の場合は,基質のある特異的な部位のユビキチン化というよりは,ユビキチン化部位を特定しない全体的な影響が大きいと考えられる.実際に,神経変性疾患の神経細胞において観察されるタンパク質の凝集体は,ユビキチンシグナルと共局在するが,より線維化しやすい直鎖型ユビキチン鎖が誘導されているケースが報告されている20).
ユビキチンをコードする四つの遺伝子のうち二つの遺伝子が,複数のユビキチンを反復する配列(直鎖型)であることから1, 2),細胞内やin vitroにおいて直鎖型ユビキチン鎖が単量体から再構築されるのは,多くのユビキチン研究者には意外な発見であったと思われる.このN末端のメチオニン基を介した直鎖型ユビキチン鎖は,それまでに非分解系タグとして同定されたK63連結型のユビキチン鎖と似た立体構造3, 21)であったことも,その生物学的意義の解明を遅らせてしまった原因の一つかもしれない.直鎖型ユビキチン鎖は,ヒトにおいて唯一のユビキチンE3リガーゼ複合体として知られるlinear ubiquitin chain assembly complex(LUBAC)により誘導される22).LUBACはheme-oxidized IRP2 ubiquitin ligase 1(HOIL-1L)/RBCK1, HOIL-1-interacting protein(HOIP)/RNF31, Shank-associated RH domain-interacting protein(SHARPIN)の三つの構成因子よりなり23–25),直鎖型ユビキチン鎖形成の活性化中心はHOIPのRBRドメインに存在する26).RBR型に属するHOIPの活性は自己抑制されており,HOIL-1LもしくはSHARPINが存在することにより,構造学的変化が誘導されて活性化される(図3a).LUBACによって誘導された直鎖型ユビキチン鎖は,先に述べたUBANドメインを持つタンパク質15, 27),さらにはA20のzinc finger(ZF)ドメインにより特異的に認識され28, 29),細胞内でシグナル複合体の形成を促す.
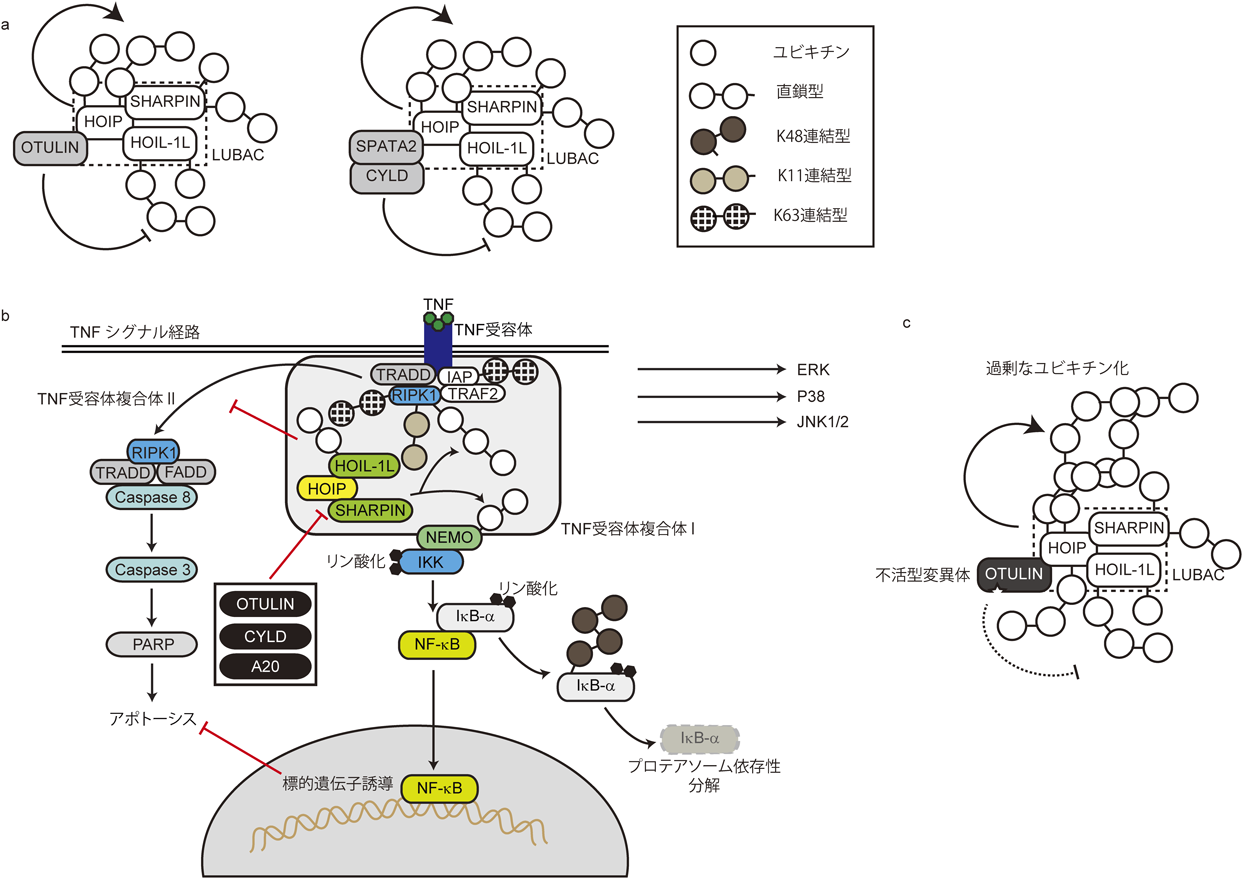
(a)直鎖型ユビキチン鎖は,LUBACを構成するHOIP, HOIL-1L, SHARPINにより誘導される.直鎖型ユビキチン鎖を加水分解するOTULINやCYLDといった脱ユビキチン化酵素はHOIPと複合体を形成し,これらにより直鎖型ユビキチン鎖は加水分解される.(b) TNF誘導性細胞内シグナルとユビキチンの役割.このシグナル経路においては,直鎖型,K11連結型,K48連結型,K63連結型のユビキチン鎖が機能する.(c) HOIPと結合するOTULINの不活型変異体は,LUBAC構成因子に結合したユビキチン鎖の加水分解を阻害することから,優性阻害型変異体として機能し,LUBAC構成因子が過剰に直鎖ユビキチン化される.
直鎖型ユビキチン鎖は,OTU deubiquitinase with linear linkage specificity(OTULIN)という直鎖型特異的な脱ユビキチン化酵素により加水分解される30, 31).興味深いことに,OTULINはHOIPと結合することが知られており(図3a,左),細胞内での直鎖型ユビキチン鎖形成の制御をHOIPとバランスをとりながら行っていると考えられる32, 33).また,cylindromatosis(CYLD)という脱ユビキチン化酵素は,SPATA2を介してHOIPと結合する(図3a,右)34–37).CYLDは直鎖型ユビキチン鎖とK63結合型ユビキチン鎖とを脱ユビキチン化する21).
このように,直鎖型ユビキチン鎖のみに着眼しても,細胞内では,その制御はダイナミックに行われ,細胞内シグナルの制御に寄与していることが想像にたやすい.
ユビキチン化が制御するあらゆる生物学的効果のうち,ここでは,多様なユビキチンコードが重要な役割を担う炎症性細胞内シグナルの制御機構について着眼する.炎症性サイトカインである腫瘍壊死因子(TNF)は下流でnuclear factor kappa B(NF-κB)経路やmitogen-activated protein kinase(MAPK)経路を活性化する(図3b)38, 39).その中でもNF-κB経路では,直鎖型,K11, K48, K63連結型の各ユビキチン鎖が重要な役割を持つことが知られる40).このシグナル経路においても,ユビキチンは,分解系(K48連結型ユビキチン鎖)および非分解系(直鎖型,K11, K63連結型ユビキチン鎖)のタグとして機能している.
TNFが結合すると,TNF受容体のDEATHドメインが細胞質においてTNFR-associated death domain(TRADD)分子をリクルートし,receptor-interacting serine/threonine-protein kinase 1(RIPK1),TNF receptor-associated factor 2(TRAF2),inhibitor of apoptosis protein(IAP)とともにTNF受容体複合体1を形成する(図3b)40, 41).RIPK1は直鎖,K11, K48, K63の各連結型のユビキチン鎖により修飾されることが知られており,これらのユビキチン化を制御する酵素も同定されている40).RIPK1は,少なくとも四つのユビキチン連結型で修飾されるが,その理由と下流シグナルの詳細な制御機構については未解明の部分が多い.ただし,RIPK1のユビキチン修飾が,ユビキチン結合ドメイン(UBD)を持つLUBACとIκB kinase(IKK)複合体をリクルートすることが,下流シグナルの活性化に作用すると考えられている.
先に述べたように,ユビキチン化は可逆的な反応で,脱ユビキチン化酵素によりユビキチン鎖は加水分解される.TNF誘導性のNF-κB活性化経路においては,脱ユビキチン化酵素活性を持つA20, OTULIN, CYLDがこの経路を負に制御する(図3b)42).これらの下流においては,転写因子NF-κBに結合し核内移行を阻害するIκB-αがIKKによりリン酸化,それに引き続くSCF β-transducin repeat containing protein(β-TrCP)によるユビキチン化によりプロテアソーム依存性に分解される43).このように,TNF誘導性のNF-κBシグナルにおいては,さまざまな酵素が異なるユビキチン型の鎖をダイナミックに制御していることがわかっている.
さて,ユビキチン化の基質へ与える影響には,ユビキチン化部位特異的なもの,および全体的なものがあると,先述した.直鎖型ユビキチンのTNF-NF-κBシグナル制御においても,この両者が関与することが示されている.全体的なものの例としては,直鎖型ユビキチン鎖特異的な脱ユビキチン化酵素OTULINの優性阻害変異体(DN-OTULIN)を発現させた細胞内では,LUBAC構成因子であるHOIPの直鎖型ユビキチン化が,異常に亢進していることがあげられる(図3c)44).これらの細胞では,制御の機序は異なるものの直鎖型ユビキチン化が起こらない細胞と同様に,TNF依存性のNF-κBシグナルの活性化は抑制される.DN-OTULINにより異常亢進したHOIPの直鎖型ユビキチン化は,LUBACのTNF受容体へのリクルートを阻害することがわかっている(図3c)44).
一方,特異的な部位のユビキチン化もNF-κBシグナルの制御に重要であることがわかってきた.直鎖型ユビキチン化の活性化中心を持つHOIPには,複数のユビキチン化部位がある.その中で,in between RING fingers(IBR)ドメインのK784に点変異を入れると,他のユビキチン化部位におけるユビキチン化や直鎖型ユビキチン鎖の形成には影響が観察されないのに対し,NF-κBの活性化は明らかに抑制される45).これらの観察結果は,HOIPのある特異的な部位にユビキチン化が起こることが,NF-κBシグナルの活性化に重要であることがわかる.さらに,TNFの下流でネクロプトーシス誘導に重要であるmixed lineage kinase domain-like(MLKL)のK219におけるユビキチン化は,その下流シグナル伝達に必要なMLKLのリン酸化や多量体の形成に重要であることも示されている46).これも,ある特異的な部位にユビキチン化が起こることによる,細胞内シグナル伝達の制御メカニズムの一つである.
ユビキチン化は,細菌と感染する宿主細胞との関係においても,さまざまな形式で関与している.サルモネラ菌を例にとってみると,少なくともLUBACとRNF213/Mysterinの二つのユビキチンリガーゼ(E3)が,感染哺乳類細胞においてサルモネラ菌のユビキチン化を誘導する(図4a)47, 48).LUBACにより形成された直鎖型ユビキチン鎖は,オートファジー受容体により認識されることによって,ゼノファジー(細菌を標的にしたオートファジー)誘導において機能する.また,サルモネラ菌感染におけるLUBACの下流では,NF-κBが活性化されることもわかっている47).サルモネラ菌表面のユビキチン化標的分子はつい最近まで不明であったが,巨大なユビキチンリガーゼRNF213が,驚いたことにエステル化によりユビキチンをリポ多糖(LPS)に結合させることが明らかにされた48).RNF213はリング(RING)ドメインを持つことから,リング型ユビキチンリガーゼとしての機能が予測されていた.興味深いことに,RNF213においては,リングドメインではなく,これまで知られていなかったジンクフィンガー(ZF)にユビキチン化の活性化中心があることが示されている(https://www.biorxiv.org/content/10.1101/2021.05.10.443411v1)48).RNF213は脳血管(内径動脈)の形態に異常をきたすもやもや病の感受性遺伝子として同定されており,これらの患者と感染制御の関連についての今後の解析も興味深いところである.これまでの研究により,LUBACとRNF213の二つのユビキチンリガーゼがサルモネラ菌のユビキチン化を誘導することが解明されたが,これら二つのユビキチンリガーゼの相互関係の詳細は不明である.
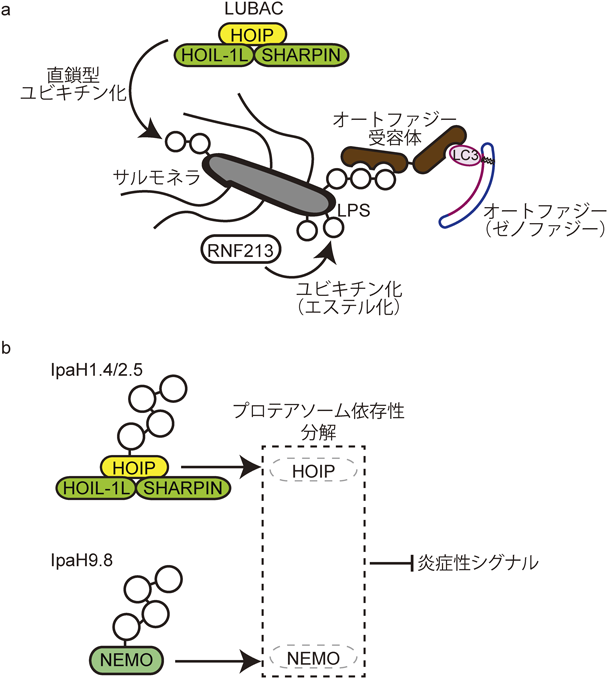
(a)サルモネラ菌が細胞に感染すると,ユビキチンにより修飾される.オートファジー受容体がこれらのユビキチン分子と結合することによって,サルモネラ菌がオートファジーによって処理される(ゼノファジー).サルモネラ菌のユビキチン化は,LUBACやRNF213によって誘導される.最近,RNF213の基質の一つがLPSであり,ユビキチンをエステル化により結合させることが明らかとなった.(b)細菌のIII型分泌装置により宿主細胞内に分泌されるユビキチンリガーゼであるIpaH1.4/2.5はHOIPを,IpaH9.8はNEMOをそれぞれユビキチン化し,プロテアソーム依存性分解を誘導することによって,炎症性シグナルを抑制する.
細菌感染と宿主細胞におけるユビキチン化としては,細菌のIII型分泌装置により宿主細胞に分泌されるユビキチンリガーゼも重要である.IpaH1.4/2.5はHOIPを49),IpaH9.8はNEMO/IKKγを50)それぞれユビキチン化し,プロテアソーム依存性の分解を誘導することによって,炎症性シグナルを抑制する(図4b).また,IpaH4.5はプロテアソームの蓋部分を構成するproteasome regulatory particle non-ATPase 13(RPN13)をユビキチン化し,プロテアソーム依存性に分解を誘導することにより,宿主細胞内のプロテアソームの機能を阻害する51).さらには,ADPリボシル化を誘導するSdeAによる宿主細胞内における内在性ユビキチン化のシステム阻害も知られる52).このように,細菌から分泌される分子によって,宿主細胞内のユビキチン化システムや標的分子に影響を与えるという機構も重要であり,宿主(ヒト)に発現する分子だけに着眼していては理解が進まないこともある.
分泌装置を介した細菌分子が新規のユビキチンコードを誘導することが明らかになったことをはじめとして,これまでに知られていなかったユビキチンコードが興味深い機構で誘導される例が,近年,次々と示されている.
ユビキチンの典型的な結合標的部位はリシン基であり,M1を介した直鎖型ユビキチン鎖は当初は新規コードと認識された.ユビキチン自身の翻訳後修飾(リン酸化やアセチル化)によるコードについても,マイトファジー53–55)やユビキチン鎖の長さの調整を行う56)ことが明らかにされてきた.ユビキチン化はイソペプチド結合(K結合型ユビキチン鎖),ペプチド結合(M1結合型ユビキチン鎖)を形成するが,最近,再度注目を集めつつあるコードは,オキシエステル結合を介したユビキチン鎖である(図5a).基本的に,最終的な基質のユビキチン化の責任を担うのはE3リガーゼであることから,E2-E3がどのように連携して,これらのコードを誘導し,基質に結合するのかを理解することが,その生物学的効果の理解にもつながると考えられる.さらには,コード特異的な脱ユビキチン化酵素の同定と制御機構の解明も重要である.エステル結合ユビキチン化については,脱ユビキチン化するエステラーゼもいくつか同定されている57).
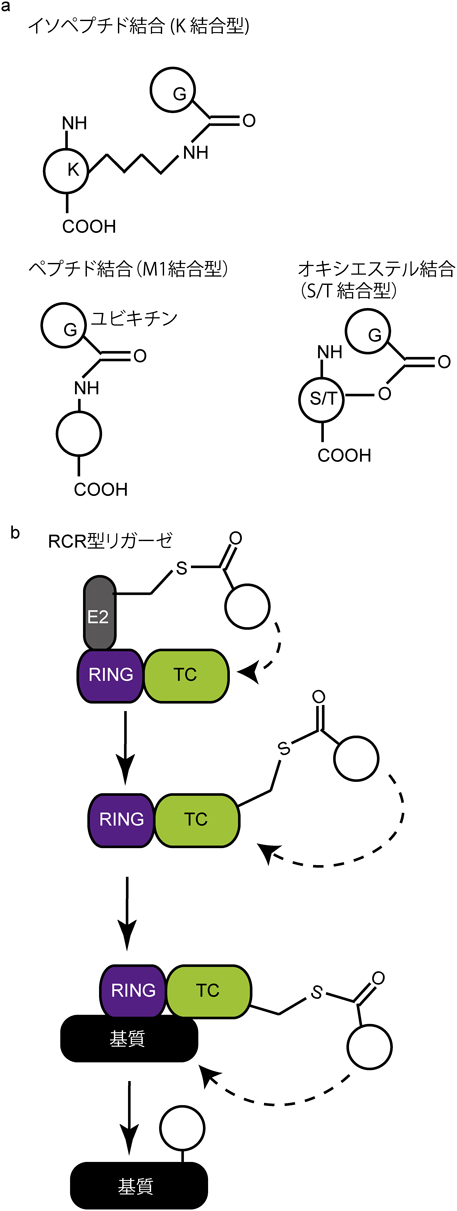
(a)ユビキチン鎖の結合型によって,イソペプチド結合(K結合型),ペプチド結合(M1結合型),オキシエステル結合(S/T結合型)を形成する.(b)新規RING-Cys-relay(RCR)型リガーゼファミリーのユビキチン化メカニズム.ユビキチンが結合した活性化E2からユビキチンがRCR型ユビキチンリガーゼのtandem cysteine(TC)ドメインに転移される.さらに,TCドメイン内でユビキチンが再度転移された後,基質がユビキチン化される.
これらの新規コードの発見は,たとえば,活性化を基盤としたプローブと質量分析を組み合わせたアプローチによるRING-Cys-relay(RCR)という新規ファミリーに属するリガーゼの同定(図5b)8)や,クライオ電子顕微鏡を用いた構造解析と生化学的解析による新規ユビキチン誘導メカニズムの解明58)などに基づく.このように,ユビキチン研究においても,技術の発展や分子ツールの開発が,その機構の理解に大きく貢献しており,技術面との連携も重要である.
筆者がユビキチン研究に携わり始めたのは,ドイツでのポスドク時代,2005年からであり,興味のあった炎症性シグナルの制御機構に,当時としては新規であったユビキチンコードの分子認識が重要であることを発見したことが現在の研究テーマ設定のきっかけであった.ユビキチンは,プロテアソーム分解の機能を持つ分子として長く知られており10),ややもすれば,古典的な研究テーマとしてとらえられるかもしれない.しかしながら,私が携わってきた15年間だけでも,さまざまな非分解系ユビキチンコード,それを操る酵素,制御の機序や制御される生物学的効果に関する多くの発見があった3–5).これらの発見に基づく知見の累積と,他分野や新規技術との連携,さまざまな分子ツールや手法の開発により,ユビキチンの研究は現在も続いて発展中である.最近では,非分解系のユビキチンの役割が特に注目されてきたようにもみえるが,proteolysis targeting chimera(PROTAC)59)やspecific and nongenetic IAP-mediated protein eraser(SNIPER)60)として知られる標的タンパク質ノックダウン法の開発により,ユビキチンの分解系タグとしての役割を最大限に活かした疾患をターゲットとする研究も大きく展開されている61, 62).
今後,明らかにされていくべきユビキチン関連の機構としては,大まかに三つのファミリーに分配されている約600のユビキチンリガーゼのより詳細な分子機構が挙げられ,その生物学的効果の理解も重要である.これまで知られていなかった機構でユビキチン化を行う分子や,多量体や複合体として機能するものもこれから同定されていくと予想される.また,ユビキチンの翻訳後修飾以外の役割,内因性のユビキチン鎖の編集の機構,新規ユビキチンコードの役割などの,ユビキチンの新規機能の理解を深めることが,さまざまな生物学的,病理学的な制御機構の解明につながることと,大きく期待される.
これまで研究を一緒に進めていただいた池田ラボのメンバー,ならびに多くの共同研究者の方々に感謝申し上げます.
1) Asaoka, T. & Ikeda, F. (2015) New insights into the role of ubiquitin networks in the regulation of antiapoptosis pathways. Int. Rev. Cell Mol. Biol., 318, 121–158.
2) Clague, M.J., Urbe, S., & Komander, D. (2019) Breaking the chains: Deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol., 20, 338–352.
3) Komander, D. & Rape, M. (2012) The ubiquitin code. Annu. Rev. Biochem., 81, 203–229.
4) Ikeda, F. & Dikic, I. (2008) Atypical ubiquitin chains: New molecular signals. ‘Protein Modifications: Beyond the Usual Suspects’ review series. EMBO Rep., 9, 536–542.
5) Swatek, K.N. & Komander, D. (2016) Ubiquitin modifications. Cell Res., 26, 399–422.
6) Wang, X., Herr, R.A., Rabelink, M., Hoeben, R.C., Wiertz, E.J., & Hansen, T.H. (2009) Ube2j2 ubiquitinates hydroxylated amino acids on ER-associated degradation substrates. J. Cell Biol., 187, 655–668.
7) Wang, X., Herr, R.A., & Hansen, T.H. (2012) Ubiquitination of substrates by esterification. Traffic, 13, 19–24.
8) Pao, K.C., Wood, N.T., Knebel, A., Rafie, K., Stanley, M., Mabbitt, P.D., Sundaramoorthy, R., Hofmann, K., van Aalten, D.M.F., & Virdee, S. (2018) Activity-based E3 ligase profiling uncovers an E3 ligase with esterification activity. Nature, 556, 381–385.
9) Kelsall, I.R., Zhang, J., Knebel, A., Arthur, J.S.C., & Cohen, P. (2019) The E3 ligase HOIL-1 catalyses ester bond formation between ubiquitin and components of the Myddosome in mammalian cells. Proc. Natl. Acad. Sci. USA, 116, 13293–13298.
10) Hershko, A. & Ciechanover, A. (1982) Mechanisms of intracellular protein breakdown. Annu. Rev. Biochem., 51, 335–364.
11) Deshaies, R.J. & Joazeiro, C.A. (2009) RING domain E3 ubiquitin ligases. Annu. Rev. Biochem., 78, 399–434.
12) Grabbe, C. & Dikic, I. (2009) Functional roles of ubiquitin-like domain (ULD) and ubiquitin-binding domain (UBD) containing proteins. Chem. Rev., 109, 1481–1494.
13) Ciechanover, A. (2005) Proteolysis: From the lysosome to ubiquitin and the proteasome. Nat. Rev. Mol. Cell Biol., 6, 79–87.
14) Dikic, I., Wakatsuki, S., & Walters, K.J. (2009) Ubiquitin-binding domains—From structures to functions. Nat. Rev. Mol. Cell Biol., 10, 659–671.
15) Wagner, S., Carpentier, I., Rogov, V., Kreike, M., Ikeda, F., Lohr, F., Wu, C.J., Ashwell, J.D., Dotsch, V., Dikic, I., et al. (2008) Ubiquitin binding mediates the NF-kappaB inhibitory potential of ABIN proteins. Oncogene, 27, 3739–3745.
16) Fennell, L.M., Rahighi, S., & Ikeda, F. (2018) Linear ubiquitin chain-binding domains. FEBS J., 285, 2746–2761.
17) Cotton, T.R. & Lechtenberg, B.C. (2020) Chain reactions: Molecular mechanisms of RBR ubiquitin ligases. Biochem. Soc. Trans., 48, 1737–1750.
18) Gundogdu, M., Tadayon, R., Salzano, G., Shaw, G.S., & Walden, H. (2021) A mechanistic review of Parkin activation. Biochim. Biophys. Acta, Gen. Subj., 1865, 129894.
19) Morimoto, D., Walinda, E., Fukada, H., Sou, Y.S., Kageyama, S., Hoshino, M., Fujii, T., Tsuchiya, H., Saeki, Y., Arita, K., et al. (2015) The unexpected role of polyubiquitin chains in the formation of fibrillar aggregates. Nat. Commun., 6, 6116.
20) Nakazawa, S., Oikawa, D., Ishii, R., Ayaki, T., Takahashi, H., Takeda, H., Ishitani, R., Kamei, K., Takeyoshi, I., Kawakami, H., et al. (2016) Linear ubiquitination is involved in the pathogenesis of optineurin-associated amyotrophic lateral sclerosis. Nat. Commun., 7, 12547.
21) Komander, D., Reyes-Turcu, F., Licchesi, J.D., Odenwaelder, P., Wilkinson, K.D., & Barford, D. (2009) Molecular discrimination of structurally equivalent Lys 63-linked and linear polyubiquitin chains. EMBO Rep., 10, 466–473.
22) Kirisako, T., Kamei, K., Murata, S., Kato, M., Fukumoto, H., Kanie, M., Sano, S., Tokunaga, F., Tanaka, K., & Iwai, K. (2006) A ubiquitin ligase complex assembles linear polyubiquitin chains. EMBO J., 25, 4877–4887.
23) Tokunaga, F., Nakagawa, T., Nakahara, M., Saeki, Y., Taniguchi, M., Sakata, S., Tanaka, K., Nakano, H., & Iwai, K. (2011) SHARPIN is a component of the NF-kappaB-activating linear ubiquitin chain assembly complex. Nature, 471, 633–636.
24) Ikeda, F., Deribe, Y.L., Skanland, S.S., Stieglitz, B., Grabbe, C., Franz-Wachtel, M., van Wijk, S.J., Goswami, P., Nagy, V., Terzic, J., et al. (2011) SHARPIN forms a linear ubiquitin ligase complex regulating NF-kappaB activity and apoptosis. Nature, 471, 637–641.
25) Gerlach, B., Cordier, S.M., Schmukle, A.C., Emmerich, C.H., Rieser, E., Haas, T.L., Webb, A.I., Rickard, J.A., Anderton, H., Wong, W.W., et al. (2011) Linear ubiquitination prevents inflammation and regulates immune signalling. Nature, 471, 591–596.
26) Stieglitz, B., Morris-Davies, A.C., Koliopoulos, M.G., Christodoulou, E., & Rittinger, K. (2012) LUBAC synthesizes linear ubiquitin chains via a thioester intermediate. EMBO Rep., 13, 840–846.
27) Rahighi, S., Ikeda, F., Kawasaki, M., Akutsu, M., Suzuki, N., Kato, R., Kensche, T., Uejima, T., Bloor, S., Komander, D., et al. (2009) Specific recognition of linear ubiquitin chains by NEMO is important for NF-kappaB activation. Cell, 136, 1098–1109.
28) Tokunaga, F., Nishimasu, H., Ishitani, R., Goto, E., Noguchi, T., Mio, K., Kamei, K., Ma, A., Iwai, K., & Nureki, O. (2012) Specific recognition of linear polyubiquitin by A20 zinc finger 7 is involved in NF-kappaB regulation. EMBO J., 31, 3856–3870.
29) Verhelst, K., Carpentier, I., Kreike, M., Meloni, L., Verstrepen, L., Kensche, T., Dikic, I., & Beyaert, R. (2012) A20 inhibits LUBAC-mediated NF-kappaB activation by binding linear polyubiquitin chains via its zinc finger 7. EMBO J., 31, 3845–3855.
30) Keusekotten, K., Elliott, P.R., Glockner, L., Fiil, B.K., Damgaard, R.B., Kulathu, Y., Wauer, T., Hospenthal, M.K., Gyrd-Hansen, M., Krappmann, D., et al. (2013) OTULIN antagonizes LUBAC signaling by specifically hydrolyzing Met1-linked polyubiquitin. Cell, 153, 1312–1326.
31) Rivkin, E., Almeida, S.M., Ceccarelli, D.F., Juang, Y.C., MacLean, T.A., Srikumar, T., Huang, H., Dunham, W.H., Fukumura, R., Xie, G., et al. (2013) The linear ubiquitin-specific deubiquitinase gumby regulates angiogenesis. Nature, 498, 318–324.
32) Elliott, P.R., Nielsen, S.V., Marco-Casanova, P., Fiil, B.K., Keusekotten, K., Mailand, N., Freund, S.M., Gyrd-Hansen, M., & Komander, D. (2014) Molecular basis and regulation of OTULIN-LUBAC interaction. Mol. Cell, 54, 335–348.
33) Schaeffer, V., Akutsu, M., Olma, M.H., Gomes, L.C., Kawasaki, M., & Dikic, I. (2014) Binding of OTULIN to the PUB domain of HOIP controls NF-kappaB signaling. Mol. Cell, 54, 349–361.
34) Wagner, S.A., Satpathy, S., Beli, P., & Choudhary, C. (2016) SPATA2 links CYLD to the TNF-alpha receptor signaling complex and modulates the receptor signaling outcomes. EMBO J., 35, 1868–1884.
35) Schlicher, L., Wissler, M., Preiss, F., Brauns-Schubert, P., Jakob, C., Dumit, V., Borner, C., Dengjel, J., & Maurer, U. (2016) SPATA2 promotes CYLD activity and regulates TNF-induced NF-kappaB signaling and cell death. EMBO Rep., 17, 1485–1497.
36) Kupka, S., De Miguel, D., Draber, P., Martino, L., Surinova, S., Rittinger, K., & Walczak, H. (2016) SPATA2-Mediated Binding of CYLD to HOIP Enables CYLD Recruitment to Signaling Complexes. Cell Rep., 16, 2271–2280.
37) Elliott, P.R., Leske, D., Hrdinka, M., Bagola, K., Fiil, B.K., McLaughlin, S.H., Wagstaff, J., Volkmar, N., Christianson, J.C., Kessler, B.M., et al. (2016) SPATA2 Links CYLD to LUBAC, Activates CYLD, and Controls LUBAC Signaling. Mol. Cell, 63, 990–1005.
38) Napetschnig, J. & Wu, H. (2013) Molecular basis of NF-kappaB signaling. Annu. Rev. Biophys., 42, 443–468.
39) Riches, D.W., Chan, E.D., & Winston, B.W. (1996) TNF-alpha-induced regulation and signalling in macrophages. Immunobiology, 195, 477–490.
40) Cockram, P.E., Kist, M., Prakash, S., Chen, S.H., Wertz, I.E., & Vucic, D. (2021) Ubiquitination in the regulation of inflammatory cell death and cancer. Cell Death Differ., 28, 591–605.
41) Annibaldi, A. & Meier, P. (2018) Checkpoints in TNF-induced cell death: Implications in inflammation and cancer. Trends Mol. Med., 24, 49–65.
42) Lork, M., Verhelst, K., & Beyaert, R. (2017) CYLD, A20 and OTULIN deubiquitinases in NF-kappaB signaling and cell death: So similar, yet so different. Cell Death Differ., 24, 1172–1183.
43) Baeuerle, P.A. & Baltimore, D. (1996) NF-kappa B: Ten years after. Cell, 87, 13–20.
44) Heger, K., Wickliffe, K.E., Ndoja, A., Zhang, J., Murthy, A., Dugger, D.L., Maltzman, A., de Sousa, E.M.F., Hung, J., Zeng, Y., et al. (2018) OTULIN limits cell death and inflammation by deubiquitinating LUBAC. Nature, 559, 120–124.
45) Fennell, L.M., Gomez Diaz, C., Deszcz, L., Kavirayani, A., Hoffmann, D., Yanagitani, K., Schleiffer, A., Mechtler, K., Hagelkruys, A., Penninger, J., et al. (2020) Site-specific ubiquitination of the E3 ligase HOIP regulates apoptosis and immune signaling. EMBO J., 39, e103303.
46) Garcia, L.R., Tenev, T., Newman, R., Haich, R.O., Liccardi, G., John, S.W., Annibaldi, A., Yu, L., Pardo, M., Young, S.N., et al. (2021) Ubiquitylation of MLKL at lysine 219 positively regulates necroptosis-induced tissue injury and pathogen clearance. Nat. Commun., 12, 3364.
47) Noad, J., von der Malsburg, A., Pathe, C., Michel, M.A., Komander, D., & Randow, F. (2017) LUBAC-synthesized linear ubiquitin chains restrict cytosol-invading bacteria by activating autophagy and NF-kappaB. Nat. Microbiol., 2, 17063.
48) Otten, E.G., Werner, E., Crespillo-Casado, A., Boyle, K.B., Dharamdasani, V., Pathe, C., Santhanam, B., & Randow, F. (2021) Ubiquitylation of lipopolysaccharide by RNF213 during bacterial infection. Nature, 594, 111–116.
49) de Jong, M.F., Liu, Z., Chen, D., & Alto, N.M. (2016) Shigella flexneri suppresses NF-kappaB activation by inhibiting linear ubiquitin chain ligation. Nat. Microbiol., 1, 16084.
50) Ashida, H., Kim, M., Schmidt-Supprian, M., Ma, A., Ogawa, M., & Sasakawa, C. (2010) A bacterial E3 ubiquitin ligase IpaH9.8 targets NEMO/IKKgamma to dampen the host NF-kappaB-mediated inflammatory response. Nat. Cell Biol., 12, 66–73, 1–9.
51) Otsubo, R., Mimuro, H., Ashida, H., Hamazaki, J., Murata, S., & Sasakawa, C. (2019) Shigella effector IpaH4.5 targets 19S regulatory particle subunit RPN13 in the 26S proteasome to dampen cytotoxic T lymphocyte activation. Cell. Microbiol., 21, e12974.
52) Bhogaraju, S., Kalayil, S., Liu, Y., Bonn, F., Colby, T., Matic, I., & Dikic, I. (2016) Phosphoribosylation of ubiquitin promotes serine ubiquitination and impairs conventional ubiquitination. Cell, 167, 1636–1649.e13.
53) Koyano, F., Okatsu, K., Kosako, H., Tamura, Y., Go, E., Kimura, M., Kimura, Y., Tsuchiya, H., Yoshihara, H., Hirokawa, T., et al. (2014) Ubiquitin is phosphorylated by PINK1 to activate parkin. Nature, 510, 162–166.
54) Kazlauskaite, A., Kondapalli, C., Gourlay, R., Campbell, D.G., Ritorto, M.S., Hofmann, K., Alessi, D.R., Knebel, A., Trost, M., & Muqit, M.M. (2014) Parkin is activated by PINK1-dependent phosphorylation of ubiquitin at Ser65. Biochem. J., 460, 127–139.
55) Kane, L.A., Lazarou, M., Fogel, A.I., Li, Y., Yamano, K., Sarraf, S.A., Banerjee, S., & Youle, R.J. (2014) PINK1 phosphorylates ubiquitin to activate Parkin E3 ubiquitin ligase activity. J. Cell Biol., 205, 143–153.
56) Ohtake, F., Saeki, Y., Sakamoto, K., Ohtake, K., Nishikawa, H., Tsuchiya, H., Ohta, T., Tanaka, K., & Kanno, J. (2015) Ubiquitin acetylation inhibits polyubiquitin chain elongation. EMBO Rep., 16, 192–201.
57) De Cesare, V., Carbajo Lopez, D., Mabbitt, P.D., Fletcher, A.J., Soetens, M., Antico, O., Wood, N.T., & Virdee, S. (2021) Deubiquitinating enzyme amino acid profiling reveals a class of ubiquitin esterases. Proc. Natl. Acad. Sci. USA, 118, e2006947118.
58) Ahel, J., Lehner, A., Vogel, A., Schleiffer, A., Meinhart, A., Haselbach, D., & Clausen, T. (2020) Moyamoya disease factor RNF213 is a giant E3 ligase with a dynein-like core and a distinct ubiquitin-transfer mechanism. eLife, 9, e56185.
59) Sakamoto, K.M., Kim, K.B., Kumagai, A., Mercurio, F., Crews, C.M., & Deshaies, R.J. (2001) Protacs: Chimeric molecules that target proteins to the Skp1-Cullin-F box complex for ubiquitination and degradation. Proc. Natl. Acad. Sci. USA, 98, 8554–8559.
60) Okuhira, K., Ohoka, N., Sai, K., Nishimaki-Mogami, T., Itoh, Y., Ishikawa, M., Hashimoto, Y., & Naito, M. (2011) Specific degradation of CRABP-II via cIAP1-mediated ubiquitylation induced by hybrid molecules that crosslink cIAP1 and the target protein. FEBS Lett., 585, 1147–1152.
61) Verma, R., Mohl, D., & Deshaies, R.J. (2020) Harnessing the power of proteolysis for targeted protein inactivation. Mol. Cell, 77, 446–460.
62) Raina, K. & Crews, C.M. (2017) Targeted protein knockdown using small molecule degraders. Curr. Opin. Chem. Biol., 39, 46–53.

九州大学生体防御医学研究所炎症制御学分野教授.博士(歯学).
1975年生.大阪で育つ.99年大阪大学歯学部卒業.2003年同大学院歯学研究科博士課程修了.研究員,特任助教を経て,05年よりゲーテ大学(ドイツ)Dikicラボに研究員として在籍.2011年よりIMBA(オーストリア)にて初めて池田ラボを主宰.19年より現職.
研究テーマと抱負炎症性細胞内シグナルの制御機構を,ユビキチン,タンパク質分解,細胞死,オートファジーに着眼し,分子レベルで解明することを目標としています.分子ツールの開発も.
ウェブサイトhttps://ikedalab.bioreg.kyushu-u.ac.jp/
趣味読書と調べもの,毎朝晩のトレーニング,時々ピアノ.

This page was created on 2021-11-02T11:58:40.218+09:00
This page was last modified on 2021-12-06T10:55:50.000+09:00
このサイトは(株)国際文献社によって運用されています。