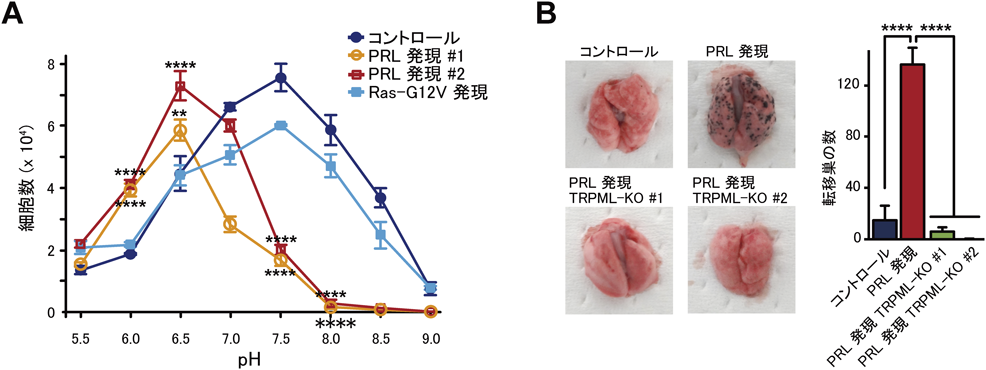
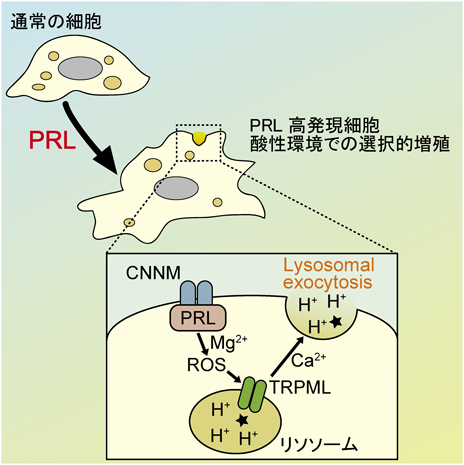
がん細胞の酸性環境への適応機構Adaptation mechanism of cancer cells to acidic environment
大阪大学微生物病研究所Research Institute for Microbial Diseases, Osaka University ◇ 〒565–0871 大阪府吹田市山田丘3–1 ◇ 3–1 Yamadaoka, Suita, Osaka 565–0871, Japan
発行日:2021年12月25日Published: December 25, 2021