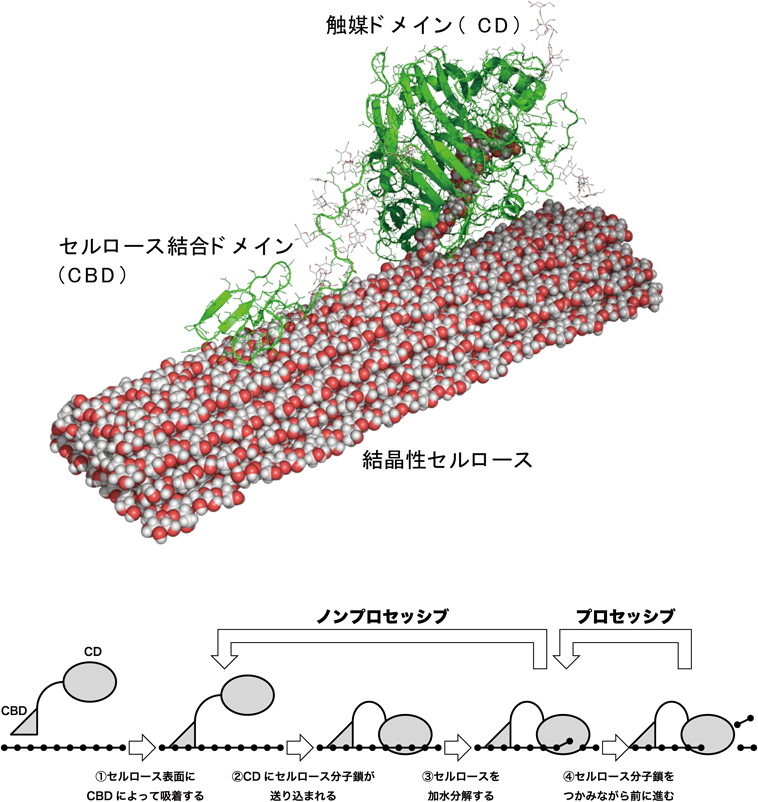
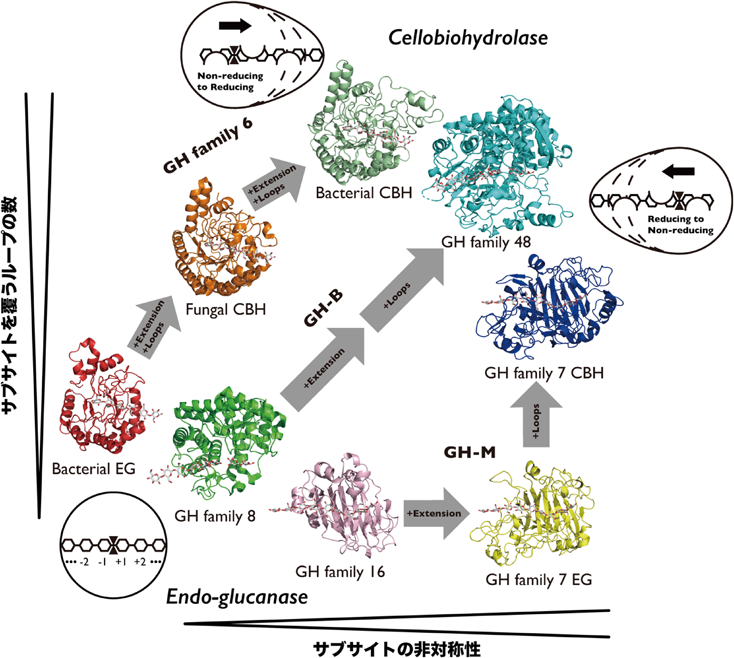
結晶性セルロースを分解するセルラーゼにおけるタンパク質レベルの収斂進化The protein level convergent evolution in cellulases degrading crystalline cellulose
東京大学大学院農学生命科学研究科Department of Biomaterial Sciences, Graduate School of Agricultural and Life Sciences, The University of Tokyo ◇ 〒113–8657 東京都文京区弥生1–1–1 ◇ 1–1–1 Yayoi, Bunkyo-ku, Tokyo 113–8657, Japan
発行日:2021年12月25日Published: December 25, 2021