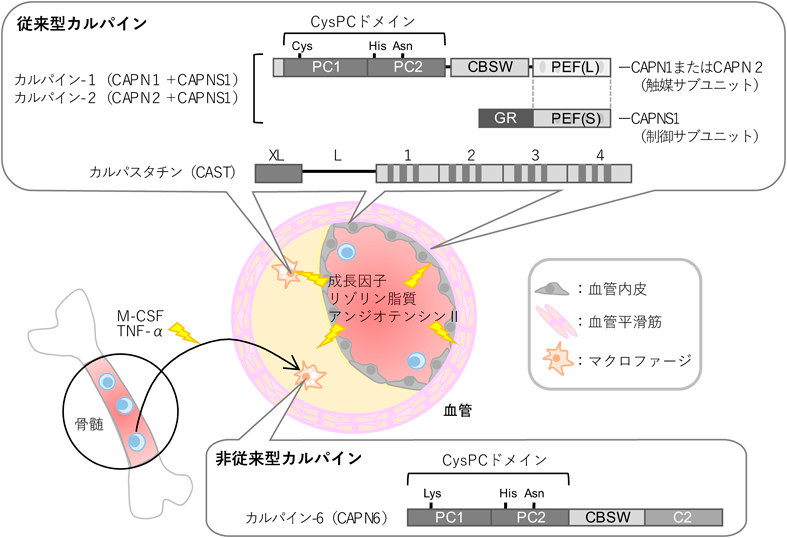
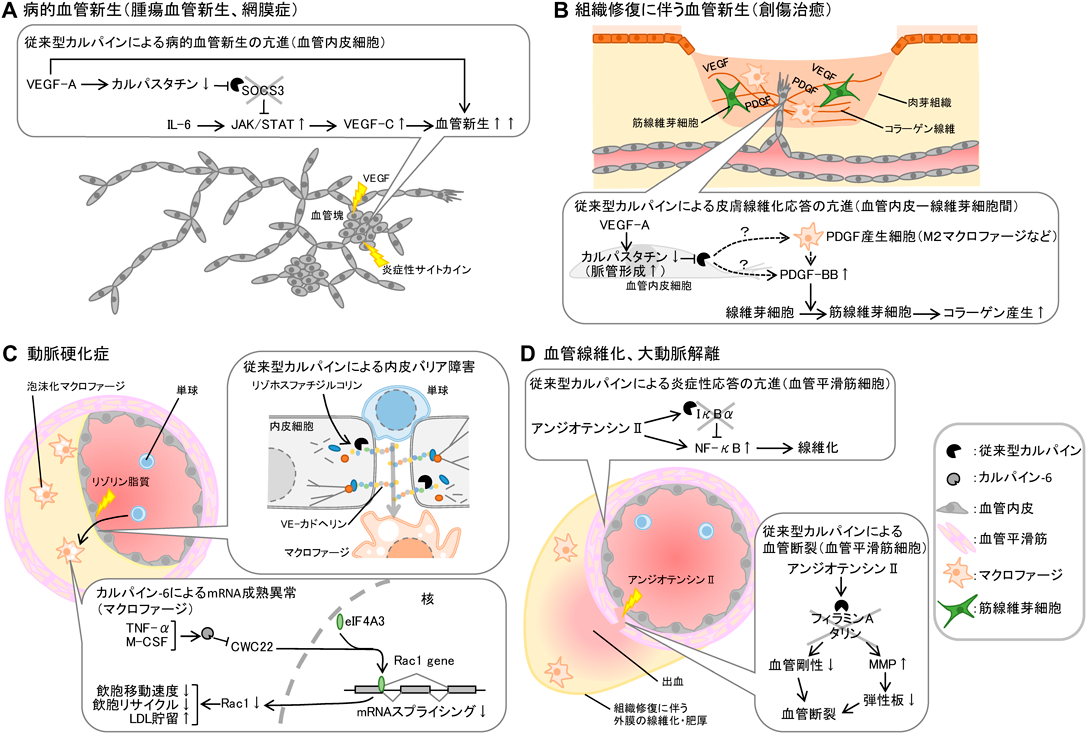
血管系疾患に潜むカルパインシステムの制御異常Pathogenic implication of vascular calpain systems
昭和大学医学部生化学講座Department of Biochemistry, SHOWA University School of Medicine ◇ 〒142–8555 東京都品川区旗の台1–5–8 ◇ 1–5–8 Hatanodai, Shinagawa-ku, Tokyo 142–8555, Japan
発行日:2022年2月25日Published: February 25, 2022