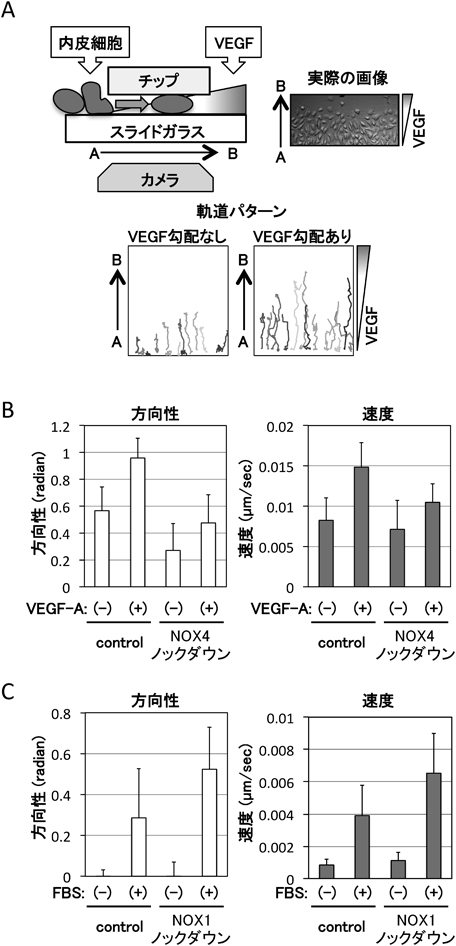
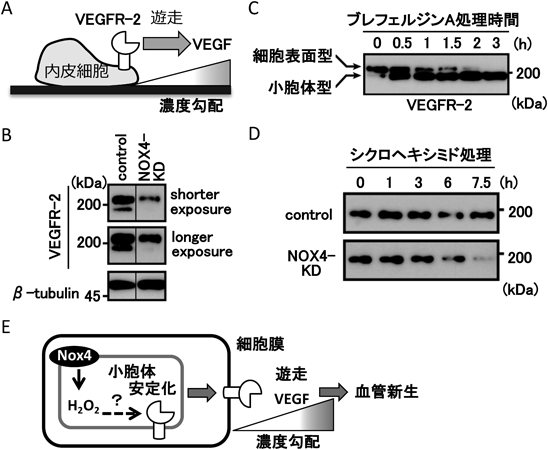
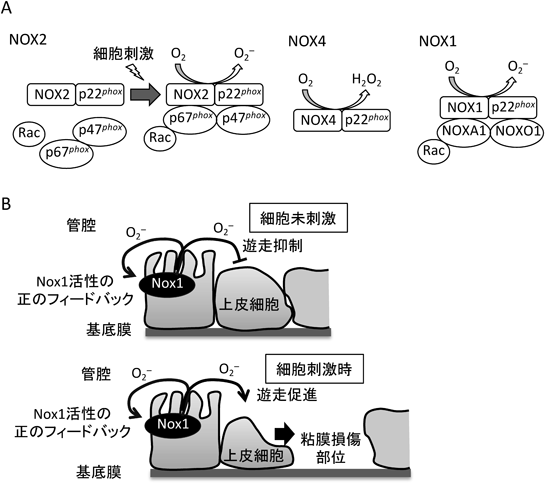
活性酸素生成酵素NOXによる細胞遊走の制御機構Regulation of cell migration by ROS-producing NOX
川崎医科大学生化学教室Department of Biochemistry, Kawasaki Medical School ◇ 〒701–0192 岡山県倉敷市松島577 ◇ 577 Matsushima Kurashiki, Okayama 701–0192, Japan
発行日:2022年2月25日Published: February 25, 2022