「MitoCarta3.0」の分類によると,代謝に関わる経路は,OXPHOSを含めて主に10カテゴリに分類できる(図1下).そのサブカテゴリにはさまざまな経路が記載されているが,いくつかの主要経路について簡単に解説する.
1)TCA(トリカルボン酸)回路
ミトコンドリアのエネルギー産生においてOXPHOSと共役する最も重要な代謝経路である.解糖系の最終産物であるピルビン酸は,ミトコンドリア内膜の輸送体(mitochondrial pyruvate carrier:MPC)を介してマトリクスに侵入するとピルビン酸デヒドロゲナーゼ複合体(PDH complex:PDC)によってアセチルCoAに変換された後,TCA回路で代謝される.この代謝系の主な機能は,①糖質,脂質,アミノ酸由来の炭素を二酸化炭素として放出すること,②OXPHOSに必要な電子をNADHとFADH2の形で供給すること,③コハク酸デヒドロゲナーゼ(または複合体II)が電子伝達系と共役していること,④アミノ酸や脂肪酸代謝のハブとして機能することである.また,TCA回路の中間体であるクエン酸,α-ケトグルタル酸,リンゴ酸は膜輸送系によってミトコンドリアと細胞質を行き来することができ,それぞれの場で代謝反応が行われていることも重要である.
2)脂肪酸のβ酸化経路
この代謝経路は,肝臓,心筋,腎臓,小腸,および褐色脂肪組織で活発に機能するとされ,逆に脳では進行しないとされている.
a.長鎖脂肪酸のミトコンドリア輸送:カルニチンシャトル
脂質が脂肪酸に分解されると,炭素数14以上の長鎖脂肪酸はミトコンドリアマトリクスのβ酸化で代謝される.この律速段階は,カルニチンシャトルと呼ばれる脂肪酸のマトリクスへの輸送段階で,それは三つの酵素反応からなる2).まず,①外膜に存在するアシルCoAシンテターゼによって脂肪酸と細胞質の補酵素A(CoA)からアシルCoAが合成された後,②同じく外膜にあるカルニチンアシルトランスフェラーゼ1(CPT1)によってアシルカルニチンに変換され,内膜のカルニチン:アシルカルニチントランスロカーゼを通過してマトリクスに流入し,③内膜のマトリクス側に位置するカルニチンアシルトランスフェラーゼII(CPT2)によってマトリクスのCoAが転移し再びアシルCoAに変換される.ちなみに炭素数12以下の脂肪酸は,カルニチンシャトルを介さずにマトリクスに入ることができる.
b.β酸化反応(飽和脂肪酸の場合)
アシルCoAは,マトリクスに存在するアシルCoAデヒドロゲナーゼ,エノイルCoAヒドラターゼ,3-ヒドロキシアシルCoAデヒドロゲナーゼ,および3-ケトアシルCoAチオラーゼの4種類の酵素によってカルボキシ末端からアセチルCoAとして2炭素ずつ酸化される.すなわち,このサイクルによって,偶数個の炭素脂肪酸を含むアシルCoAはすべてアセチルCoAに変換される.この代謝系の重要な機能は,①TCA回路にアセチルCoAを供給すること,②OXPHOSに必要な電子をNADHとFADH2の形で供給することである.なお,奇数個の炭素脂肪酸を含むアシルCoAは,β酸化の最終過程でプロピオニルCoA(炭素数3)を生じ,その後,三つの反応を経てTCA回路に組み込まれる.すなわちプロピオニルCoAは,①ビオチンを必要とするプロピオニルCoAカルボキシラーゼによってメチルマロニルCoAに変換され,②メチルマロニルCoAエピメラーゼによって可逆的にエピマー化され,③最終的にビタミンB12(コバラミン)を必要とするメチルマロニルCoAムターゼによって可逆的に,TCA回路中間体のスクシニルCoAに変換される.
c.β酸化反応(不飽和脂肪酸の場合)
天然に存在する不飽和脂肪酸は,9位や12位にシス二重結合を持つため,上記経路の反応に伴って2位と3位にシス二重結合を持つ中間体が生成する.これらは,NADPHを補酵素とする2,4-ジエノイルCoAレダクターゼ(DECR1)による還元作用を受けトランス体となった後,イソメラーゼの作用で2-トランス体に変換され通常のβ酸化系に入る.
3)ケトン体合成
肝臓や腎臓のミトコンドリアにおいて,アセチルCoAが過剰となった場合にケトン体が合成される.ケトン体とは,アセト酢酸,β-ヒドロキシ酪酸,アセトンのことを指す.この反応は,飢餓(またはインスリン不足)時にエネルギー源としての糖質が低下し脂質代謝が亢進した場合,OXPHOSに障害が起きた場合(ミトコンドリア障害)やTCA回路がうまく機能しないときに進行するとされる.まず,2分子のアセチルCoAがチオラーゼによって縮合しアセトアセチルCoAに変換された後,さらにアセチルCoAと縮合して,HMG-CoA(β-ヒドロキシ-β-メチルグルタリルCoA)が生じる.その後,開裂してアセト酢酸が生成し,D-β-ヒドロキシ酪酸デヒドロゲナーゼによる可逆反応によって3-ヒドロキシ酪酸が生じる.一方,アセト酢酸はアセト酢酸デカルボキシラーゼによって脱炭酸してアセトンに変換される.これらの反応で生じたケトン体は肝臓では代謝されず,血流を介して骨格筋,心筋,腎皮質,脳に運ばれ,そこでアセチルCoAに代謝されTCA回路に入る.すなわち,ケトン体は肝臓以外の組織ではエネルギー源として利用される(特に脳では糖に代わる重要なエネルギー源となる).
4)ステロイド合成
ステロイドホルモンは副腎皮質や生殖腺(精巣,卵巣,胎盤)において臓器特異的に合成される.これらの生合成はすべてコレステロールを原材料とするが,律速となる最初の反応がミトコンドリア内膜(のマトリクス側)で起きる.実際,ステロイド合成に特化した副腎では,ミトコンドリアが豊富に存在し,内膜の面積を稼ぐためクリステが高度に発達している.反応はまず,細胞質からミトコンドリアへ遊離コレステロールが輸送され,内膜のCYP11A1(シトクロムP-450ファミリー)によって側鎖が切断されプレグネノロンを生成する.プレグネノロンはすべてのステロイドホルモンの前駆体であり,小胞体(ER)に輸送された後,それぞれ特異的な反応を受け各種ホルモンが合成される.
5)カルジオリピン
カルジオリピン(CL)は,二つのリン酸基と4種類の脂肪酸鎖を有するミトコンドリア特異的なリン脂質であり,肝臓,心臓,腎臓のミトコンドリア内膜の約20%を占めるとされる.生合成の最初のステップでは,ERにおいてCDP-ジアシルグリセロール合成酵素(CDP-DAG synthase)によってホスファチジン酸(PA)とCDPが連結し,CDP-DAGを生成する.CDP-DAGがミトコンドリアに輸送されると,内膜の①ホスファチジルグリセロリン酸合成酵素(PGPS)によってホスファチジルグリセロリン酸(PGP)となり,②PGPホスファターゼによってホスファチジルグリセロール(PG)を生成する.さらに③カルジオリピン合成酵素(CLS)によってPGとCDP-DAGが縮合することでCLが生成される.その後,④CLはリモデリング(再アシル化反応)によって不飽和度の高い脂肪酸に置換されて成熟する.タファジン(TAZ)は心臓や骨格筋のミトコンドリアで発現が強くみられ,CLのリモデリングに関与している.
6)ユビキノン代謝
ユビキノン(またはコエンザイムQ)はミトコンドリア内膜に存在し,電子伝達系において必須の役割を担う3).ユビキノンはキノン骨格を有する4-ヒドロキシ安息香酸(4-HB)とイソプレン鎖を前駆体とする.4-HBはチロシンに由来し,イソプレン鎖は細胞質のメバロン酸経路に由来する.ヒトはイソプレン単位が10個のCoQ10が最も多いが,CoQ9もCoQ8も一部存在する(マウスではCoQ9が最も多い).ユビキノンの生合成経路は内膜あるいは内膜のマトリクス側で起きるとされる.4-HBとイソプレン鎖が独立にマトリクスに輸送された後,PDSS1/PDSS2によってイソプレン鎖が連結し,次にCOQ2によって4-HBとイソプレンポリマーが連結する.その後,COQ酵素群による修飾反応を受け成熟する4).実際,ユビキノンはミトコンドリア以外の膜画分(たとえばER,ミクロソーム,ゴルジなど)においても確認できるが,その輸送経路は不明である.また,ユビキノンは複合体IとIIに加えて,他の内膜局在型の酸化還元酵素,たとえば電子伝達フラビンタンパク質デヒドロゲナーゼ(ETFQOR),グリセロールリン酸デヒドロゲナーゼ(GPDH),ジヒドロオロト酸デヒドロゲナーゼ(DHODH),コリンデヒドロゲナーゼ(CHDH),プロリンデヒドロゲナーゼ(PRODH),硫化物:キノンオキシドレダクターゼ(SQOR)などからも電子を授受することができる.
7)鉄代謝
生命活動に必須の金属元素である鉄は,細胞質とミトコンドリアのそれぞれにおいてバランスを一定に保つ鉄恒常性の仕組みが働いていると考えられる.ミトコンドリアでは,鉄輸送(import/export)に関わるトランスポーター群,鉄を貯蔵するフェリチン(FTMT),ヘム生合成経路,および鉄–硫黄クラスター(Iron–Sulfur cluster:ISC)タンパク質群が関与するとされる5).特に,電子伝達系の複合体IからIVは,ヘムとISCのいずれか(複合体IIおよびIIIは両方)を含むため,鉄代謝はOXPHOSの機能維持に必須といえる.
a.ポルフィリン生合成:ヘム合成
ポルフィリンは四つのピロール環が閉環した環状化合物であり,その誘導体であるプロトポルフィリンの生合成は生体にとってきわめて重要である.ポルフィリン環の中に配位する金属によって機能が異なる.鉄(Fe)を配位したものがヘムであり,代表的なヘムタンパク質は電子伝達に関わるシトクロムb(複合体IIIの構成タンパク質),シトクロムc,ヘモグロビン,ミオグロビン,カタラーゼ,一酸化窒素合成酵素(NOS),P-450ファミリーなどである.一方,コバルト(Co)を配位したものがビタミンB12(シアノコバラミン)である.ヘムの生合成において,第1反応と後半の第6, 7反応,およびヘム合成(第8反応)の4種類の反応がミトコンドリアで行われる(ポルフィリンの生合成は第7反応まで).すなわち,①マトリクスにおいて,グリシンとスクシニルCoAがδ-アミノレブリン酸シンターゼ(ALAS1)によってδ-アミノレブリン酸(5-ALA)を生成する.この反応はプロトポルフィリン,およびヘム合成の律速段階とされ,ヘムによるフィードバック制御を受ける.5-ALAは細胞質に移行して,いくつかの反応を経た後,コプロポルフィリノーゲンIIIがミトコンドリアに再移行し,②内膜に局在するコプロポルフィリノーゲンIIIオキシダーゼ(CPOX)による脱炭酸反応によってプロトポルフィリノーゲンを生成し,③プロトポルフィリノーゲンオキシダーゼ(PPOX)によってプロトポルフィリンを生成する.④ヘム生合成の最終反応は内膜のマトリクス側に局在する鉄付加酵素(FECH)によって,プロトポルフィリンに鉄(Fe2+)が付加される.
b.Fe–Sクラスター生合成
真核生物において,ミトコンドリアはISC生合成の場として考えられている6).硫黄もまた生体に必須の微量元素であり,鉄は硫黄原子もしくはタンパク質のシステイン側鎖の硫黄原子と会合する.ISCを含むタンパク質としては,電子伝達系の複合体I, II, III, TCA回路のアコニターゼ,フェレドキシン,リポ酸合成酵素(LASY)などがあげられる.ISC生合成に関わる酵素は複合体を形成し,その構成タンパク質は生物種でよく保存されているとされる.主な構成因子はピリドキサール5-リン酸(PLP)を補酵素とするシステインデスルフラーゼ(NFS1)とその安定化タンパク質(ISD11, ACP1),フラタキシン(FXN),およびスキャフォールドタンパク質(ISCU)などが知られている.
8)アミノ酸代謝
アミノ酸は,内膜に存在するトランスポーター群によってマトリクスに移行し,脱アミノ化を受けた後はさまざまな代謝反応を受け,最終的にTCA回路に入るか,生体成分に組み込まれるかのいずれかの運命をたどる.
a.分岐鎖アミノ酸
分岐鎖アミノ酸(BCAA)のバリン,ロイシン,イソロイシンは必須アミノ酸であり,異化経路はほぼすべてミトコンドリアに存在する.最初の2ステップは共通しており,まず①分岐鎖アミノ酸脱アミノ基転移酵素(BCAT)によって脱アミノ化反応を受けた後,②分岐鎖ケト酸脱水素酵素複合体(BCKDH)によって不可逆的に脱炭酸反応を受ける.この反応が律速段階であるとともに,特異的なキナーゼ(BDK)とホスファターゼ(BDP)によるリン酸化調節(リン酸化されると不活性化)を受けるとされる.この結果,バリンはイソブチリルCoAとなり,いくつかの反応を経た後,スクシニルCoAとなってTCA回路に入る.一方,ロイシンとイソロイシンはそれぞれイソバレリルCoAと2-メチルブチリルCoAになり,これらは最終的にアセチルCoAとなってTCA回路に入る.
b.グルタミン酸
グルタミンは,グルタミナーゼ(GS,グルタミン合成酵素)によってグルタミン酸とアンモニアに変換される.この反応は可逆的であり,脳,腎臓,肝臓に存在し,特にアストロサイトでは神経毒のアンモニアと興奮毒のグルタミン酸をグルタミンに変換して血中へ放出するとされる.グルタミン酸はその後,グルタミン酸デヒドロゲナーゼ(GDH),後述するアスパラギン酸アミノ基転移酵素(mAST),およびアラニンアミノ基転移酵素(mALT)のいずれかによってα-ケトグルタル酸となり,TCA回路に入る.
c.グリシン
マトリクスのグリシンは,上述したように,①ヘム合成の第1反応において重要な基質となる.また②肝臓,腎臓,脳,胎盤の内膜に局在する4種類の酵素系であるグリシン開裂酵素群(P, H, T, Lタンパク質)によって,後述するテトラヒドロ葉酸(THF)とともに基質となって,二酸化炭素とアンモニア,5,10-メチレンテトラヒドロ葉酸(5,10-CH2-THF)に分解される.グリシンの炭素は5,10-CH2-THFのメチレン骨格に組み込まれる.
d.セリンと葉酸代謝回路(1炭素代謝)
ミトコンドリアにおけるセリンの代謝は,ミトコンドリアと細胞質にまたがる葉酸代謝回路に組み込まれる形で進行する7).この反応の意義は,①NADH/NADPHを供給すること,②ミトコンドリアのタンパク質合成反応に関与することの2点である.食事から摂取された葉酸は,細胞質のNADPH要求性のジヒドロ葉酸還元酵素(DHFR)によってTHFとなる.THFは内膜の輸送体(mitochondrial folate carrier, SLC25A32)によってマトリクスに移行し,セリンとともにセリンヒドロキシメチル転移酵素(SHMT2)の基質となり,可逆的にグリシンと5,10-CH2-THFに変換される.5,10-CH2-THFはtRNAのアンチコドン1字目のタウリノメチルウリジン修飾(τm5U)の供与体として重要であり,このタウリン修飾によってコドン3字目の認識が甘くなる,いわゆるゆらぎ対合(wobble base-pair)を生み出し,1種類のtRNAが複数のコドンを解読することを可能としている8).この反応にはMTO1とGTPBP3が関与するとされる.5,10-CH2-THFはさらにNAD+/NADP+要求性のメチレンテトラヒドロ葉酸デヒドロゲナーゼ(MTHFD2)によって10-ホルミルテトラヒドロ葉酸(10-formyl-THF)に変換される.その後10-formyl-THFは三つの運命をたどる.①真核生物のミトコンドリアにおいて,メチオニルtRNAホルミルトランスフェラーゼ(MTFMT)によってtRNAMetのメチオニンにホルミル基が付与される(生成物としてTHFが再生する).これがミトコンドリア遺伝子の翻訳開始コドンにロードされ,タンパク質合成が始まる9).②10-ホルミルテトロヒドロ葉酸デヒドロゲナーゼ(mtFDHまたはALDH1L2)によってTHFとCO2に変換される.あるいは③MTHFD1LによってTHFとギ酸に変換される.THFとギ酸は細胞質に移行した後,同様の代謝反応を受け,プリンやチミジンなどの核酸前駆体,あるいはメチオニンの生合成に利用される.
e.尿素回路
アンモニアは上記のGSやGDHによる代謝反応で生成するが,逆に同化する酵素はマトリクスに存在するカルバモイルリン酸合成酵素(CPS1),GS,およびGDHの三つである.哺乳動物は最終的にアンモニアを尿素として排出するが,この代謝経路に関係する酵素はすべて肝臓に存在する.一部の反応がミトコンドリアマトリクスを介しており,①CPS1はこの回路の律速段階で,不可逆的にアンモニアを取り込みカルバモイルリン酸に変換する.この反応は,N-アセチルグルタミン(NAG)によって促進する.さらに②オルニチンカルバモイルトランスフェラーゼ(OTC1)によってカルバモイルリン酸とオルニチンが縮合しシトルリンを生成する.その後,シトルリンは細胞質へ移行した後,さらに3段階の反応を受け,尿素が生成する.
9)糖新生
糖新生は,低血糖に応答したグルカゴンによって発動する代謝反応であり,主に肝臓と腎臓で行われ,一部の反応はミトコンドリアのTCA回路を経由する.糖新生基質は乳酸,ピルビン酸,アミノ酸,プロピオン酸である.乳酸はピルビン酸に変換され,ピルビン酸はマトリクスのピルビン酸カルボキシラーゼ(PC)によってオキサロ酢酸に変換される.この反応が糖新生の律速段階と考えられている.オキサロ酢酸はリンゴ酸デヒドロゲナーゼ(MDH2)によってリンゴ酸になり,後述するリンゴ酸–アスパラギン酸シャトルを経由して細胞質に移行し,細胞質のリンゴ酸デヒドロゲナーゼ(MDH1)によって再びオキサロ酢酸になり,ホスホエノールピルビン酸カルボキシキナーゼ(PCK)によってホスホエノールピルビン酸に変換された後,解糖系を逆行する.アミノ酸はトランスアミナーゼによって脱アミノ化された後,ピルビン酸,α-ケトグルタル酸,スクシニルCoA,フマル酸,オキサロ酢酸のいずれかに変換され,最終的にはオキサロ酢酸から糖新生経路に入る.
10)ミトコンドリアシャトル系
解糖系で生成したNADHは,ミトコンドリア内膜を透過できない.そこで,NADHに含まれる電子をミトコンドリアに輸送するため,特別なシャトル経路が存在する7).
a.リンゴ酸–アスパラギン酸シャトル
肝臓,腎臓,心臓のミトコンドリアで活発に機能するNADHシャトル系であり,ミトコンドリアと細胞質に存在する六つの酵素反応からなる.すなわち,細胞質のNADHは①リンゴ酸デヒドロゲナーゼ(MDH1)によってオキサロ酢酸に電子を渡してリンゴ酸とNAD+を生成する.リンゴ酸は,②内膜のリンゴ酸–α-ケトグルタル酸アンチポーターによってマトリクスに入り,③リンゴ酸デヒドロゲナーゼ(MDH2)によってNAD+に電子を渡してNADHとオキサロ酢酸を生成する.このシャトルを駆動するため,オキサロ酢酸はさらに④アスパラギン酸アミノ基転移酵素(mAST)によってアスパラギン酸をグルタミン酸とα-ケトグルタル酸に変換した後,②および⑤グルタミン酸–アスパラギン酸アンチポーターによってグルタミン酸とα-ケトグルタル酸が細胞質に輸送され,最後に⑥細胞質のASTによってオキサロ酢酸とアスパラギン酸が再生する.
b.グリセロールリン酸シャトル
骨格筋や脳,褐色脂肪組織で利用されているシャトル系である.細胞質のNADHの電子は,①グリセロールリン酸デヒドロゲナーゼ(cGPDH)によってジヒドロキシアセトンリン酸に電子を渡してグリセロールリン酸を生成する.さらに②ミトコンドリア内膜のグリセロールリン酸デヒドロゲナーゼ(mGPDH)によって電子がFADからFADH2を介してユビキノンに伝達される.このシャトル系は,ミトコンドリアのNAD+/NADHに直接影響を及ぼさない点が上記のリンゴ酸–アスパラギン酸シャトル系と異なっている.
11)ヌクレオチド生合成
ヌクレオチドの生合成は,新生(de novo)経路と再生(salvage)経路があり,増殖が活発な細胞は新生経路を優先的に利用し,筋肉や脳など非増殖細胞では再生経路が使われているとされる10).ミトコンドリアDNAやRNA合成に必要なヌクレオチドは,増殖細胞ではほぼ細胞質から供給されている.ピリミジンヌクレオチドとプリンヌクレオチドの新生経路の合成過程は異なっているが,一つの酵素を除き,すべての反応が細胞質で行われている.
a.de novo経路
ピリミジンヌクレオチド新生経路の4番目の反応はミトコンドリアで行われる.中間体であるジヒドロオロト酸は,ミトコンドリア内膜の膜間腔側に局在するジヒドロオロト酸デヒドロゲナーゼ(DHODH)によってオロト酸に変換され,再び細胞質で代謝される.DHODHはフラビンモノヌクレオチド(FMN)を有しており,ジヒドロオロト酸から電子を受け取ってFMNH2を生成し,さらに内膜のユビキノンに電子を受け渡している.
b.salvage経路
ミトコンドリアマトリクスには,ヌクレオチドの分解産物であるヌクレオシドを再生する酵素として,デオキシグアノシンキナーゼ(DGUOK)とチミジンキナーゼ(TK2)の2種類のサルベージ酵素が存在する.また,モノリン酸ヌクレオチドキナーゼ群(NMPKs)やジリン酸ヌクレオチドキナーゼ群(NDPKs)も存在する.特に,DGUOKはミトコンドリアのプリンデオキシリボヌクレオチド(dNTP)合成の律速酵素と考えられている.
12)アルコール代謝
飲酒すると,エタノールの80%は小腸で,20%は胃から吸収される.その後,血流を介して主に肝臓(他に脳,膵臓,胃)で代謝される.まず細胞質のアルコールデヒドロゲナーゼ(ADH)によってアセトアルデヒドが生成し,その後ミトコンドリアのアルデヒドデヒドロゲナーゼ(ALDH2)によって酢酸に分解され,酢酸はアセチルCoAとなってTCA回路に入る.
13)活性酸素種の生成とその消去系
活性酸素種(reactive oxygen species:ROS)は,酸素に由来する細胞障害性の高い分子の総称で,スーパーオキサイドアニオン(·O2−),過酸化水素およびヒドロキシラジカル(·OH)などが知られている(このうち·O2−と·OHは不対電子を持つことからフリーラジカルとも呼ばれる).ミトコンドリアは生体の約95%の酸素を消費するとされるが,このうちの0.1~4%は電子伝達系から漏れ出た電子還元によって·O2−が生成し,複合体Iのフラビン部位やISR-ユビキノン部位,複合体IIのフラビン部位,複合体IIIのユビキノール酸化部位(Qo site/center P)が主要な発生源とされている11).マトリクスの·O2−はスーパーオキシドジスムターゼ2(SOD2/Mn-SOD)によって水と過酸化水素に分解される.複合体IIIから発生した·O2−は,マトリクスだけでなく膜間腔にも放出されるが,こちらはSOD1(Cu/Zn-SOD)やαトコフェロール(ビタミンE)などによって除去される.過酸化水素はタンパク質や核酸などの生体高分子に対して強力な酸化剤として働く一方で,細胞外に移行しシグナル分子として働く役割も知られている.さらに,過酸化水素はFe2+やCu+を介したフェントン反応によって,より細胞障害性の高い·OHに変換される.このため,マトリクスには過酸化水素を速やかに処理する二つの消去系が備わっており,NADPHの供給がこの仕組みを維持するうえで不可欠な役割を果たしている.
a.PRX-TRX回路
この回路は,通常のミトコンドリア呼吸によって生じた生理的レベル(数nM程度)の過酸化水素を除去する上で主要な役割を果たしている.マトリクスのペルオキシレドキシン3(PRX3)はチオレドキシン(TRX2)を電子供与体として利用することで過酸化水素を水に還元する.チオレドキシン還元酵素2(TXNRD2)は,NADPHを利用してPRX3/TPX2の還元状態を維持する役割を果たしている.
b.GSH/GPX経路
グルタチオン(glutathione:GSH)を介した反応系はより高濃度(いわゆる酸化ストレスレベル)の過酸化水素(または過酸化脂質など)に対して働く防御システムと考えられている.GSHは三つのアミノ酸からなるトリペプチド(γ-グルタミル–システイニル–グリシン)で,細胞内の主要な抗酸化物質として機能する.主に細胞質で合成された後,全体の10~15%がミトコンドリアに存在するとされる.最近,GSHの輸送に必要な内膜のトランスポーターSLC25A39(とそのパラログであるSLC25A40)が同定された12).GSHペルオキシダーゼ(GPX1)は2分子のGSHから電子を奪って過酸化水素を水に還元する.酸化型グルタチオン(GSSG)は,NADPHに依存したグルタチオン還元酵素(GR)によってGSHにリサイクルされる.GSSGは内膜を通過できないため,GRの活性がミトコンドリアのGSSGレベルを調節している11).
14)NAD+代謝
ミトコンドリアの代謝は,OXPHOSを含めて電子の授受を伴う酸化還元反応が多い.NAD+やNADP+はさまざまなデヒドロゲナーゼの補酵素として,またNADHやNADPHは還元酵素の電子供与体としてきわめて重要な役割を果たしているが,このうち,マトリクスの内膜を通過できるのは唯一NAD+のみである(細胞質のNADHは,上述したシャトル系を介して還元当量がマトリクスに伝わる).最近,哺乳類細胞において,NAD+の輸送に必要な内膜のトランスポーターSLC25A51(とそのパラログであるSLC25A52)が同定された13).従来,マトリクスのNAD+は,前駆体であるニコチンアミドモノヌクレオチド(β-NMN)からNMNAT3によって生合成されると考えられていたが,現在では否定されている14).すなわち,ミトコンドリアのNAD+は細胞質からの輸送によってレベルが規定される.また,マトリクスのNAD+の一部はNADキナーゼ(NADK2)によってNADP+に変換される15).すなわち,NADP+のレベルはこのキナーゼ活性に依存して規定されている16).
5. 代謝産物の網羅的解析法としてのメタボローム解析
生体の代謝産物とは,分子量2000未満の小分子化合物(アミノ酸,核酸構成成分,炭水化物,ペプチドなど)と,脂肪酸やリン脂質などの脂溶性分子を含めたものを指し,メタボローム解析は,質量分析MSなどによって代謝産物を網羅的に計測する手法のことである.従来の細胞抽出液を用いたメタボローム解析は,細胞に含まれるすべての代謝産物を対象とする.上述したように,ミトコンドリアを場とする多くの代謝産物はミトコンドリアにとどまっているわけではなく,細胞質や他のオルガネラと相互に行き来する.したがって,従来の細胞メタボローム解析では,ミトコンドリア内の代謝動態を推測することは基本的に不可能であると考えられる.そこで我々は,ミトコンドリアを標的とした新しい解析法「ミトコンドリア-メタボロミクス」を開発した19).
1)ミトコンドリア単離法の改良
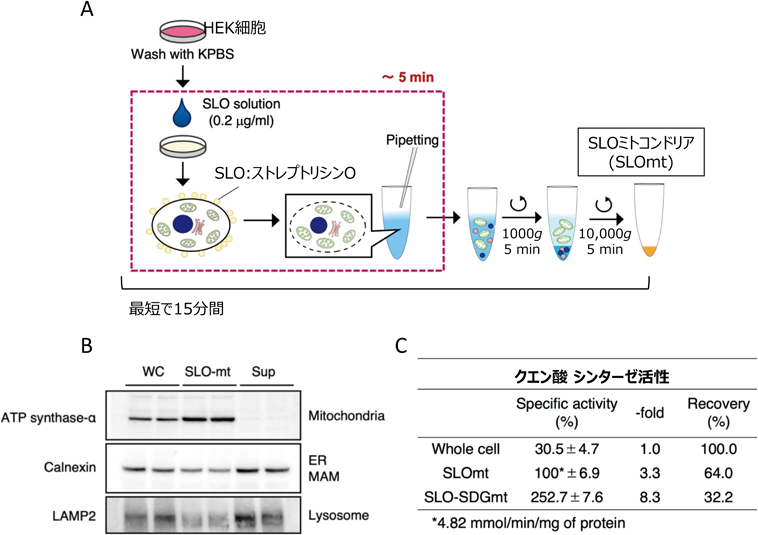
従来から行われてきたミトコンドリア単離法は,まず細胞を氷冷下に置き,ダウンス型やポッター型のホモジナイザーを使って破砕し,その後,低速遠心法によって核や未破砕の細胞を沈殿させた後,中速遠心分離によってミトコンドリア膜画分を得た.ショ糖密度勾配(sucrose density gradient:SDG)遠心分離をさらに行うことで,より精製度の高いミトコンドリア画分を得ることができる.しかし,このような処理は時間と労力がかかり,精製の過程でミトコンドリアの代謝物が変化してしまうリスクがあった.そのため,より迅速にミトコンドリアを単離し,できるだけ代謝変化を抑えることができる手法が求められた.そこで以前報告された,溶血性レンサ球菌(Streptococcus pyogenes)の溶血毒素であるストレプトリシンO(SLO)を用いた穏やかなミトコンドリア単離法20)を改良し,メタボローム解析用に最適化を行った.このワークフロー(図2A)によって,HEK293細胞からわずか15分の間に濃縮されたミトコンドリア画分(以下,SLOmt)を得ることができる.しかも,同時に多検体を処理することができるので,ホモジナイザーを使って一つずつ処理する手間を省くことができる.SLOmtタンパク質については,ウエスタンブロット解析の結果,複合体Vのサブユニットタンパク質(ATP合成酵素α)が実質的に濃縮されており(図2B),クエン酸シンターゼCSの比活性は細胞抽出液に比べて約3.3倍高い値を示した(図2C).
2)SLOmtからの代謝物抽出,メタボローム解析
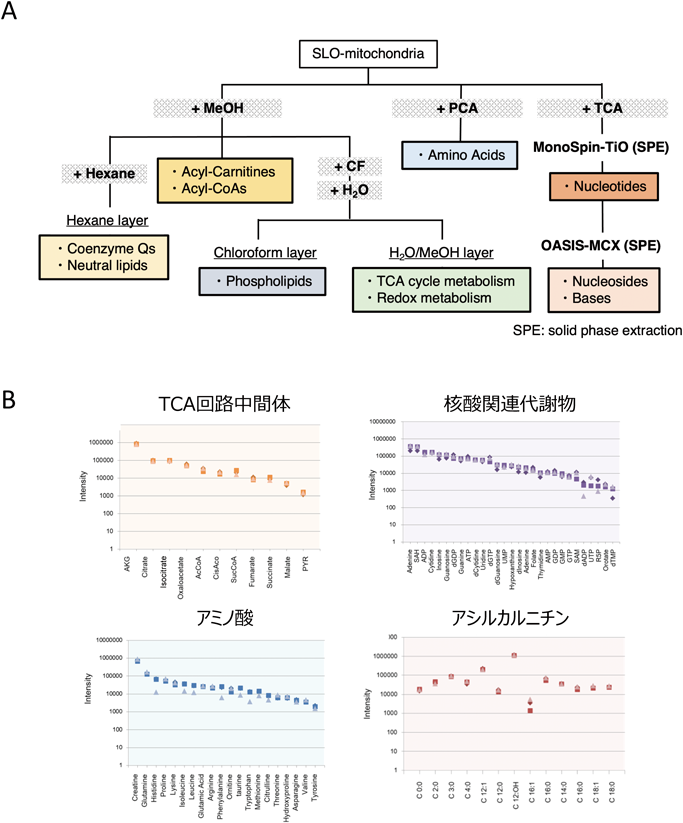
上述したように,ミトコンドリアには多くの代謝酵素が局在するため,その代謝産物も多種多様含まれていると考えられる.我々は,その特性に応じた抽出法を確立(図3A)し,液体クロマトグラフィー質量分析(LCMS)による独自メソッドを構築した.この技術開発によって,SLOmtに含まれる代謝産物(たとえばTCA回路中間体,ヌクレオチド,アミノ酸,アシルカルニチン,リン脂質,コエンザイムQなど)を数百種類以上測定することが可能となった(図3B).
3)ミトコンドリアの精製度と代謝物
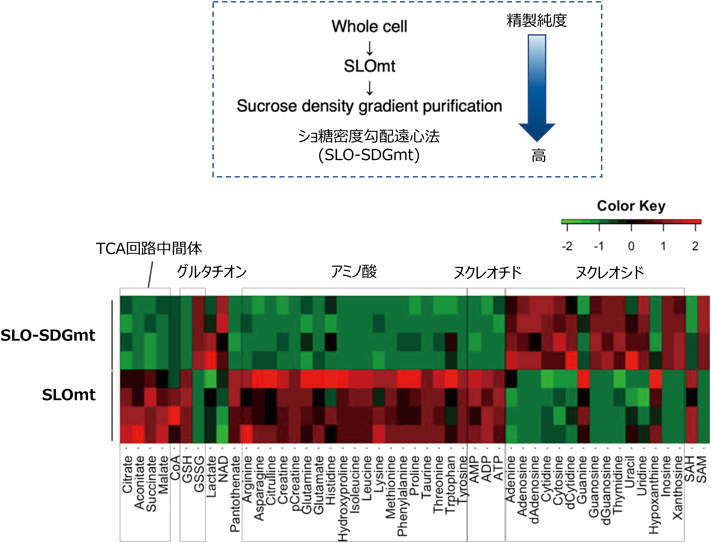
従来の生化学的な考え方では,ミトコンドリアの精製度を上げるとミトコンドリアに含まれる成分の純度が上がるとされる.実際,SLOmtをショ糖密度勾配遠心によってさらに精製したSLO-SDGmtは,CSの比活性でみるとSLOmtの2倍以上濃縮されていた(図2C).そこで,代謝産物もタンパク質と同様に濃縮されるかどうかを検証するため,単位タンパク質あたりの代謝物を比較した.その結果,TCA回路の中間体(クエン酸やコハク酸など)やエネルギー代謝物(ATPなど)は,SLO-SDGmtで相対的に減少していた(図4).また,ヌクレオチドの分解産物であるヌクレオシドや塩基は上昇した.これらの結果は,SLO-SDGmtの代謝産物は精製に時間がかかるため,その過程に異化反応(ヌクレオチド分解など)が進んでしまった可能性が考えられた.ミトコンドリアに豊富に存在し抗酸化作用を示すグルタチオン(GSH)は,SLO-SDGmtでは主に酸化型(GSSG)が多く存在することがわかった.GSSGは活性酸素種(ROS)の指標であるため,超遠心処理過程において酸化ストレスが発生した可能性も示唆される.
4)代謝物の急速な変化を捉えるSLOmtの精度評価
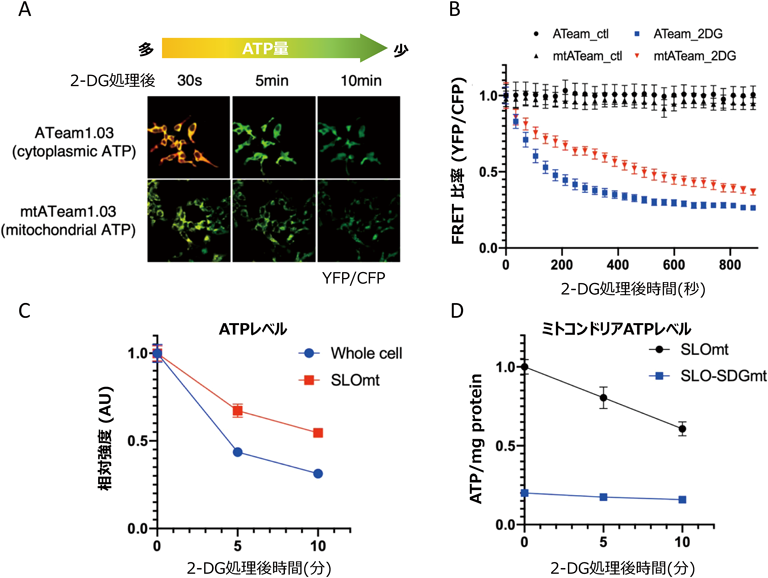
本手法がミトコンドリア内で起きる急速な代謝変化を捉えることができるかどうか,についてさらに検討した.そのため,今村博士らが開発したATeam 1.03と呼ばれるFRETプローブを用いて,解糖系阻害した際の細胞内およびミトコンドリア内のATPをライブで観察した(図5A).細胞質ATPは,阻害剤処理後10分以内に急速に減少したのに対し,ミトコンドリアATPは直線的な減少を示した(図5B).この処理時間の下で,5分後と10分後の細胞からSLOmtを精製すると,そのATPの相対強度は,顕微鏡で観察した変化率とほぼ一致していた(図5C).また,SLO-SDGmtのATPシグナルは低いレベルにとどまっており,定量的な変化を検出することはできなかった(図5D).この結果からも,SLO-SDGmtはメタボローム解析用途に適していないことが再確認された.