ヒストンバリアントの形成するクロマチン構造と機能Structure and function of chromatin containing histone variants
公益財団法人がん研究会がん研究所The Cancer Institute of JFCR ◇ 〒135–0063 東京都江東区有明3–8–31 ◇ 3–8–31 Ariake, Koto-ku, Tokyo 135–0063, Japan
発行日:2022年4月25日Published: April 25, 2022
ヒストンバリアントは主要型ヒストンの亜種であり,ヒストンの化学修飾やDNAメチル化と同様にクロマチンがDNAの安定維持および遺伝子の発現制御に機能するために必須の因子である.精巣をはじめとする組織特異的に発現するヒストンバリアントが同定されており,精巣特異的なヒストンバリアントの欠失は精子形成が行われず雄性不妊の原因となる.さらに,ヒストンバリアントの変異が細胞のがん化の原因となることも報告されている.このようにヒストンバリアントは真核生物にとってきわめて重要な因子である.本稿では,ヒストンバリアントが形成するクロマチンの構造と機能およびヒストンバリアントが特定のクロマチンに取り込まれる機構について解説する.
© 2022 公益社団法人日本生化学会© 2022 The Japanese Biochemical Society
ゲノムDNAは遺伝情報を持つ生命にとって必須な物質である.ゲノムDNA中にコードされている遺伝子から適切なタイミングで適切な量のタンパク質が作られることで細胞は機能している.この転写はエピジェネティクスと呼ばれるDNA配列によらない機構で制御されている.真核生物の場合は,エピジェネティクス機構はゲノムDNAが形成するクロマチン構造が鍵となっている.本稿ではクロマチン構造の基本構成因子であるヒストンの亜種「ヒストンバリアント」が,エピジェネティクス機構においてどのように機能しているかについて解説する.
真核生物のゲノムDNAは,クロマチン構造を形成して細胞の核内に収納されている.クロマチン構造の基本単位は,ヌクレオソームと呼ばれる4種類のヒストンタンパク質(H2A, H2B, H3, H4)各2分子からなるヒストン八量体に約150塩基対のDNAが巻きついた構造体である1).クロマチンは,DNA上で起きる反応である転写,複製,組換え等を制御する重要な構造体である.
細胞学的な研究から,クロマチンは転写が活発に行われているユークロマチンと転写が抑制されているヘテロクロマチンに分かれることが明らかとなった2).後の研究より,ユークロマチン領域はDNase Iヌクレアーゼに対してヘテロクロマチン領域より感受性が高いことから,ユークロマチンは凝集度が低く,ヘテロクロマチンは凝集度が高いことが示された3).これらのことから,クロマチン構造の違いにより遺伝子の転写が変化することが明らかとなった.クロマチン構造は発生や分化により細胞の形質が転換する際に可逆的に変換される.さらに,近年のクロマチン相互作用解析技術の発展により,ユークロマチンおよびヘテロクロマチンは一様な構造を形成しているのではなく,トポロジカルドメイン(topologically associating domain:TAD)という相互作用頻度の高いひとつながりの領域を形成していることが明らかとなった4).トポロジカルドメインの変化は遺伝子の転写制御において重要である.たとえば,ユークロマチンとヘテロクロマチンのトポロジカルドメインが隣り合っている領域において,細胞の形質転換に伴いヘテロクロマチンのトポロジカルドメインが大きくなり,ユークロマチン内で転写されていた遺伝子がヘテロクロマチンのトポロジカルドメイン内に局在することで転写が抑制されるといったことが観察されている.このようにクロマチン構造が柔軟に変化することは遺伝子の転写制御においてきわめて重要である.
一方で,細胞の形質転換や細胞分裂を経ても一定のクロマチン構造を形成している領域が存在する.分裂期染色体の一次狭窄部位であるセントロメアや染色体の末端領域であるテロメアである.セントロメアは分裂期に紡錘糸が姉妹染色分体に結合するキネトコアが集積する領域であり,染色体の均等分配に必須の領域である.テロメアは,DNA末端からヌクレアーゼによって削られないように保護するために重要であり,細胞の老化やがん化と密接に関わる.これらの領域は,その機能不全は細胞死につながるため,きわめて安定に維持されている.
このような多様なクロマチンの構造と機能は,DNA配列によらないエピジェネティックな機構で制御されている.これまでにエピゲノム情報と呼ばれるヒストンの化学修飾,ヒストンバリアント,DNAメチル化がクロマチンの構造や機能と相関することが明らかになっている.これらはクロマチンの基本単位であるヌクレオソームの構成因子であり,ヌクレオソームレベルでのバリエーションがクロマチンの構造および機能を決定している.実際に,エピゲノム情報に特異的に結合するクロマチン因子が存在し,同じDNA上の領域においてもエピゲノム情報が異なることでクロマチンに結合する因子が変化する.このことが多様なクロマチンの構造と機能につながる.本稿では,エピゲノム情報の一つであるヒストンバリアントの形成するクロマチンの構造と機能について解説する.
ヒストンバリアントは主要型ヒストン(H2A, H2B, H3, H4)と比べて相同性が高いが,別の遺伝子にコードされている亜種である.ヒストンバリアントの多くはH2AおよびH3のバリアントであるが,ヒストン八量体がH2A–H2B複合体とH3–H4複合体からなることを考えると,それぞれの複合体にバリアントを含むバリエーションがあることがわかる.
主要型ヒストンはS期特異的に発現し,DNA複製と共役しヌクレオソームを形成する(図1).そのため,ゲノムDNA全体にわたり局在する.ヒストンバリアントの多くは細胞周期に依存せず発現しており,DNA複製非依存的にヌクレオソームを形成する.この反応は既存のヌクレオソーム中のヒストン複合体と置き換わる反応であるため,ヒストン交換反応(histone exchange, histone turnover)と呼ばれる(図1).ヒストン交換反応はゲノム全体にわたって同じ頻度で起きるわけではなく,各ヒストンバリアントにそれぞれ交換反応の頻度の高いゲノムDNA上の領域が存在する.この性質により,ヒストンバリアントの局在は限定される.ヒストン交換反応は,クロマチンに取り込まれる前のヒストン複合体に結合する因子が担っており,これらヒストンに結合し,クロマチンへの取り込みを行うタンパク質をヒストンシャペロンと呼ぶ.ヒストンバリアント特異的なヒストンシャペロンが同定されており,ヒストンシャペロンは相同性が高い主要型ヒストンとヒストンバリアントを区別して結合していることが構造解析や生化学的な解析から明らかとなっている.また,クロマチンに取り込まれる前のヒストンはヒストンシャペロン以外にもさまざまな因子と相互作用しており,これらの相互作用因子も主要型ヒストンとヒストンバリアントの間で異なる.そのため,クロマチンに取り込まれる前のヒストンバリアント複合体もヒストンバリアントの機能を考える上で重要である.これ以降,研究の進んでいるヒストンバリアントを中心に解説する.
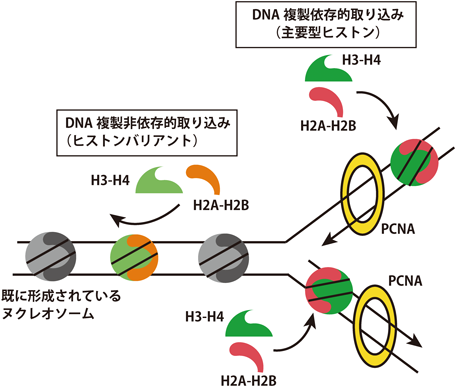
ヒストンがクロマチンに取り込まれるにはDNA複製に依存した機構(右)と依存しない機構(左)が存在する.主要型のヒストンおよび一部のヒストンバリアント(H2A.Xなど)はDNA複製依存的にクロマチンに取り込まれるが,多くのヒストンバリアントはDNA複製非依存的にクロマチンに取り込まれる.
S期に複製された姉妹染色分体は,M期に染色体を形成して,二つの娘細胞に均等に分配される.染色体の均等分配時に両極から伸びてきた紡錘糸が結合するキネトコア複合体が形成されるゲノムDNA上の領域がセントロメア領域であり,M期染色体では一次狭窄部位となる.ヒトの場合,セントロメア領域はアルフォイドDNAと呼ばれる反復配列からなるため,このDNA配列がセントロメア領域を規定していると考えられた.しかし,ネオセントロメアというアルフォイドDNA上ではない領域に機能的なセントロメアが安定に維持されている事例が発見されたことから,アルフォイドDNAはセントロメアを規定する因子ではないことがわかった.セントロメア領域にはヒストンH3のバリアントであるCENP-Aをはじめとするセントロメア領域に特異的に局在するタンパク質群が存在する5).著者および他のグループの実験により,CENP-AをゲノムDNA上のセントロメア領域とは異なる領域に局在させたところ,他のセントロメアタンパク質が集積することが明らかとなった6–8).このことから,ヒストンH3バリアントであるCENP-Aがエピジェネティックにセントロメア領域を規定していることがわかった.また,CENP-Aは酵母,線虫,ショウジョウバエ,ニワトリ,マウス,ヒトなどの生物種において広く保存されており,これらの生物種において遺伝学的な研究よりCENP-Aはセントロメア領域の形成および維持に必須の因子であることが明らかとなっている.
CENP-Aヌクレオソームがセントロメア領域を規定する機構として,CENP-Aヌクレオソームの構造が主要型のH3ヌクレオソームと異なり,その構造がエピジェネティックにセントロメア領域の形成および維持に機能していることが考えられた.著者のグループは2011年にX線結晶構造解析法によりヒトのCENP-Aヌクレオソームの構造を明らかにした9).その結果,CENP-Aヌクレオソームはヒストン複合体と安定的に結合しているDNAが短く,両端のDNAの配向が通常のヌクレオソームとは異なり決まっていないことがわかった.ヌクレオソームの両端のDNAは隣のヌクレオソームにつながっていることから,このCENP-Aヌクレオソームの性質はセントロメア領域の高次クロマチン構造に影響を与えることが考えられた.そこで,我々はCENP-Aヌクレオソームの両端に主要型のH3ヌクレオソームが形成しているトリ・ヌクレオソームを試験管内において再構成を行い,クライオ電子顕微鏡を用いて解析を行った10).その結果,CENP-Aヌクレオソームはトリ・ヌクレオソームの構造を変換させることがわかり,セントロメア領域のクロマチン構造はCENP-Aヌクレオソームにより特殊な構造を形成することがわかった.さらに,CENP-Aに特異的にみられるループ構造は,セントロメアタンパク質であるCENP-Nが結合する領域であり,CENP-AのC末端領域にはCENP-Cが結合することがわかった11, 12).これらのことから,CENP-Aヌクレオソーム自身が他のセントロメアタンパク質が集積するための基盤構造となっていることがわかる.
このようにCENP-Aはセントロメア領域の形成および維持に必須であるため,CENP-Aが細胞分裂を経てもセントロメア領域にヌクレオソームを形成する機構を明らかにすることが,セントロメア領域が安定的に維持されるメカニズムを理解する上で重要である.これまでにCENP-Aがセントロメア領域に取り込まれるのは,分裂期の後期からG1期の初期に起こることが明らかとなっている13, 14).CENP-Aのクロマチンへの取り込みは,CENP-A特異的なヒストンシャペロンHJURPが必要である13, 14).HJURPはクロマチンに取り込まれる前のCENP-A-H4複合体に結合しており,分裂期にセントロメア領域に局在するMis18複合体と相互作用することで,CENP-Aのセントロメア領域への集積に機能していることが考えられる(図2)15).さらに,HJURPはセントロメアタンパク質の一つであるCENP-Cともクロマチンを介さずに結合する6).CENP-Cは自身で多量体化する性質があるため,HJURPに結合したCENP-Cとセントロメア領域に局在するCENP-Cが相互作用することが,HJURPのセントロメア領域の認識に機能しているかもしれない.
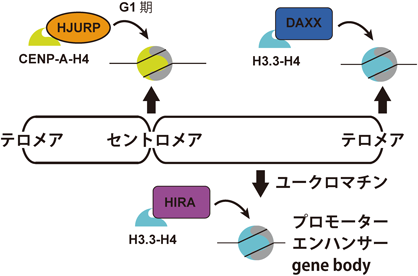
CENP-AはHJURP依存的にセントロメア領域のクロマチンに取り込まれる.また,染色体の脱凝縮が終わったG1期に取り込まれる.H3.3はHIRA依存的にユークロマチン中の転写が行われているプロモーター,gene bodyおよびエンハンサーに取り込まれる.また,テロメア領域や近傍のヘテロクロマチン領域にはDAXX依存的に取り込まれる.
CENP-Aの過剰発現は多くのがん細胞で報告されている.培養細胞を用いた実験から,過剰発現したCENP-Aはセントロメア領域以外のクロマチンにも取り込まれることが観察された16).CENP-Aの異所局在は染色体上に複数のセントロメア領域が形成され,染色体の均等分配がうまくいかず,染色体不安定性(chromosome instability:CIN)を引き起こす可能性がある.実際にCENP-Aの欠失や過剰発現は,いずれも染色体の構造および数の異常であるCINを促進する.さらに,CENP-Aのタンパク質の量の多さは,がん細胞の侵襲性の増加と相関していることが報告されている.また,CENP-Aの過剰発現はがん抑制遺伝子であるp53が正常に機能しているかどうかで細胞の形質に与える影響が変わる17).p53が正常であれば,CENP-Aの過剰発現は老化と放射線感受性を促進する.しかし,p53を不活性化している細胞においてCENP-Aが過剰発現すると,がん細胞の浸潤や転移につながる上皮間葉転換を促進する.このように,CENP-Aは染色体の均等分配だけではなく細胞の形質獲得にも機能することが明らかとなっている.
精巣では精原細胞が分化して精子が形成される.精子のクロマチンは大部分のヒストンタンパク質が塩基性タンパク質であるプロタミンに置き換わり,精子形成の過程において大規模なクロマチンの再編成が行われる.さまざまな生物種における精子や精巣のヒストンの初期の研究により,雄の生殖細胞でのみ発現するヒストンバリアントの存在が明らかになった.これらのヒストンバリアントは,プロタミンへの置き換わりに先行して,主要型ヒストンと置き換わっている.そのため,精巣特異的なヒストンはプロタミンへの置き換わりに必要だと考えられる.
精巣特異的なH3バリアントであるH3tは,精原細胞の分化に伴い徐々に発現し,主要型のH3と置き換わる(図3)18).ノックアウトマウスの解析から,雄のマウスにおいてH3tの欠損は精子が形成されず完全な不妊となる18).H3tノックアウトマウスでは,減数分裂した細胞が少なかったことから,H3tが減数分裂への移行に関与している可能性が示唆された.また,雌のマウスではH3tの発現が確認されず,ノックアウトマウスにも特段の表現型も観察されない.マウスのH3tを含むヌクレオソームは主要型H3を含むヌクレオソームと比べて,両端のDNAとヒストン複合体との結合が弱く,H3tヌクレオソームからなるポリヌクレオソームは凝集しにくい性質を持つ.これは,H3tの42番目のアミノ酸がヒスチジンである(主要型H3ではアルギニン)ことが原因である.H3 R42はヌクレオソームのDNAと相互作用しているが,H3t H42ではその相互作用が弱まっている.また,同様にヒトのH3tを含むヌクレオソームにおいても,主要型H3を含むヌクレオソームと比べてきわめて安定性が低いことが生化学的解析から明らかとなっている19).これらのことから,H3tの形成するヌクレオソームが不安定であることが,精原細胞から分化する際に機能している可能性が考えられる.
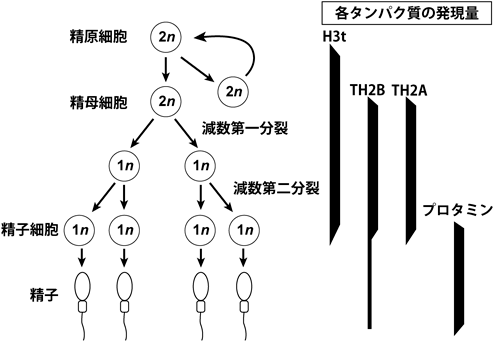
精子では多くのヒストンがクロマチンから取り除かれ,プロタミンと呼ばれる塩基性のタンパク質に置き換わっている.この分化の過程において,精巣特異的なヒストンバリアントが発現し,主要型のヒストンと置き換わることでプロタミンへの置換を促進している.
H3tに加え,精巣特異的なH2BバリアントであるTH2Bが同定されている20).H2BのバリアントはH2AおよびH3に比べて種類が少なく,精巣特異的なバリアントが存在することは興味深い.TH2Bは減数分裂開始とともにクロマチン中のH2Bと置き換わり,大部分がTH2Bのヌクレオソームになる(図3).しかし,TH2Bが欠失していても精子形成は正常に行われ,雄の生殖能力に影響を与えない.TH2Bノックアウトマウスでは,通常型のH2Bの発現量が増えることでTH2Bの欠損を補っている.さらに,野生型では観察されないヒストンの化学修飾がTH2Bの欠損したクロマチンでは観察される21).同定されたこれらの化学修飾はDNAと結合するリシン残基のクロトニル化(H3 K122とH4 K77)もしくはアルギニンのメチル化(H 4R35, H4 R55, H4 R67およびH2B R72)であり,ヌクレオソームの安定性を弱めることが示唆される.したがって,TH2Bの欠損は,正常なH2Bの過剰発現だけでなく,ヌクレオソームを不安定にする効果を持つヒストン化学修飾により相補されていることが示唆される.これらのことから,TH2Bの主な機能は,ゲノムスケールで安定性の低いヌクレオソームを形成し,プロタミンとの置き換わりに機能していることが考えられる.実際に,X線結晶構造解析および生化学的解析によって,TH2BおよびTH2A(TH2Bと共発現するH2A変異体)を含むヌクレオソームは,ヒストンとDNAの相互作用が著しく減少し,ヒストン–ヒストン相互作用が変化することで,ヌクレオソームの安定性にさらに劇的な効果があることが示された22).H3tと異なりTH2BおよびTH2Aは卵母細胞においても発現しており,TH2B-TH2Aを含むヌクレオソームの安定性の低さが卵の形成過程においても機能していることが考えられる.さらに興味深いことに,TH2B, TH2AおよびヒストンシャペロンであるNPM2を体細胞に発現させると,リプログラミング因子(Oct4, Sox2, Klf4, c-Myc:OSKM)によるiPS細胞化(人工多能性幹細胞化)を促進することが報告された.これはTH2BおよびTH2Aがよりアクセスしやすいクロマチンを形成するためだと考えられる23).
転写が活性化されているのか,抑制されているのかは,その遺伝子領域に存在するヒストンの化学修飾の種類と相関する.転写が活性化している領域には,H3 K4me3, H3 K14ac, H3 K27acなどの修飾が存在しており,一方で転写が不活性化されている領域ではH3 K9me3, H3 K27me3などが存在している.これらの化学修飾と同様にヒストンバリアントの局在も転写状態によって変化することが知られている.
H3バリアントであるH3.3は複製非依存的にクロマチン中に取り込まれる.ほとんどの真核生物は,複製依存的にクロマチンに取り込まれる主要型のH3と複製に依存しないH3.3バリアントを発現しているが,菌類など一部の真核生物はH3.3タイプのみを発現している24).H3.3はH3とわずか4もしくは5個のアミノ酸が異なるだけであり,これらのアミノ酸のうち三つは,コアヒストンフォールドドメイン内にみられる25, 26).H3.3を含むヌクレオソームのクロマチン中の存在量は,分裂している細胞と終末分化した細胞で異なる.終末分化した神経細胞では,H3.3ヌクレオソームは全H3の約90%を占めるが27),分裂中の細胞では,H3.3は10~20%である28, 29).これは,終末分化した細胞ではDNAが複製しないため新たなH3.1の取り込みは起こらずに,H3.3との交換反応が進むためであると考えられる.また,増殖している細胞ではH3.3の取り込みは,転写が活性化している遺伝子プロモーター,gene body領域,エンハンサーなどの領域で起きる30).これらのことを併せて考えると,H3.3は,これらの領域を認識して集積しているのではなく,これらの領域がヒストンの交換反応頻度が高い結果としてH3.3の取り込みが顕著に観察されていることが考えられる.一方で,哺乳類の胚性幹細胞(embryonic stem cells:ESC)において,転写が抑制されておりヒストンの交換反応の頻度が低い遺伝子上にH3.3の濃縮が報告されている31).これらのことから,ヒストンの交換頻度に依存したH3.3の取り込みに加え,H3.3のクロマチンへの取り込みに関する明らかとなっていない機構が存在することが示唆される.H3.3は転写活性化と抑制の両方に機能する潜在性を持ち,おそらくは化学修飾やクロマチン結合因子などの組合わせによって,転写を制御していると考えられる.
H3.3のクロマチンへの取り込みは,転写活性化および不活性化の両方の領域においてhistone regulatory factor A(HIRA)複合体に依存している(図2)32).さらに,HIRAに依存しないメカニズムも見つかっている.death-associated protein(DAXX)はH3.3特異的シャペロンとして同定され,SWI/SNFファミリーリモデラーであるα-thalassemia/mental retardation syndrome protein(ATRX)とともに,テロメアやその周辺領域のヘテロクロマチンへのH3.3の取り込みを行う(図2)31, 33).なぜ,領域によってヒストンシャペロンが使い分けられているかは明らかではないが,ヒストンの交換頻度が低く反応性が乏しいヘテロクロマチンへH3.3を取り込むために,ATRXなどのATP依存的なクロマチンリモデリング活性が必要なのではないかと考えられる.
H3.3の変異体が特定のがん細胞においてがん化の原因となっている(ドライバー変異)ことが報告された.H3.3 K27M(27番目のリシンがメチオニンに変異)が小児のグリオーマにおいて,H3.3 K36M(36番目のリシンがメチオニンに変異)が軟骨肉腫と骨巨細胞腫においてドライバー変異体である.H3のK27やK36はメチル化修飾を受け,クロマチンの機能に関与している.これらのアミノ酸がメチオニンに変異することで,特定のリシンメチル化酵素がメチル化することができなくなりエピジェネティックな遺伝子発現制御が破綻することががん化の原因と考えられる.しかし,これらの変異が特定の組織においてのみがん化のドライバーとなる理由は明らかとなっていない.
H2A.Zは広く保存されているH2Aバリアントである.H2A.Zと転写の関係は,テトラヒメナの研究から最初に報告された.テトラヒメナには転写活性の高い大核と低い小核があるが,H2A.Zは大核のクロマチンにのみ取り込まれていることから,H2A.Zは転写活性化に機能していることが考えられた34).さまざまな細胞種や生物においてH2A.Zのゲノム上での局在を解析した結果から,H2A.Zは主にプロモーターやエンハンサーなどの制御領域に限定されていることがわかり,これらの領域に存在するヒストン化学修飾と共局在する.著者らは培養細胞を界面活性剤で処理し,細胞膜と核膜に孔を開けた透過性細胞のクロマチンを鋳型としてH2A.Zの取り込みを調べた35).その結果,H2A.Zは転写が活性化している遺伝子のプロモーターおよびエンハンサーに取り込まれること,H2A.Zはランダムにクロマチンに取り込まれた後にこれらの領域に取り込まれた分子以外がクロマチンから取り除かれるのではなく,これらの領域を標的としてクロマチンに取り込まれることがわかった.また,転写が活性化している遺伝子は細胞種によって異なるため,転写が活性化しているプロモーターおよびエンハンサーに存在するヒストン化学修飾やクロマチン結合因子が,H2A.Zが取り込まれる領域の目印となっていることが考えられる.実際に,クロマチンに取り込まれる前のH2A.Z複合体にはヒストンアセチル化に結合するYEATSドメインを持つGAS41が含まれる36).GAS41はH3のK18およびK27がアセチル化されたペプチドに特異的に結合する.これらの化学修飾は転写活性化領域に存在する.GAS41はC末端のドメインで二量体化するため,2分子のGAS41がそれぞれアセチル化リシンと結合することで安定した結合となる.ただし,H3K18acとK27acの局在は必ずしも一致しないため,細胞内ではこの他の組合わせ(H3 K27acとH4のリシンアセチル化など)で結合していることも考えられる.さらに,クロマチンに取り込まれる前のH2A.Z複合体にはヒストンアセチル化酵素であるTIP60(KAT5)も含まれる.そのためH2A.Zが転写活性化しているプロモーターおよびエンハンサーを標的にすることで,ヒストンアセチル化酵素がこれらの領域に集積し,ヒストンのアセチル化レベルが保たれている可能性がある.
一方で,H2A.Zヌクレオソームは転写抑制のマークであるH3 K27me3が含まれることが共免疫沈降実験で示されている37).これらのことから,H2A.Zは転写活性化と抑制の相反する機能を有していることが示唆される.現在では,H2A.Zは,化学修飾やH2A.Z以外のバリアントがヌクレオソームに含まれるかなどを含む複数の要因によって,転写の活性化および抑制化に関与することが明らかになっている.
H2AのC末端の領域はヌクレオソームの末端のDNAと相互作用し,DNAとヒストン複合体の結合を安定させる1).H2A.BバリアントはC末端の領域が欠失しており,DNAとヒストン複合体の結合が弱いヌクレオソームを形成する38).HeLa細胞におけるH2A.Bのノックダウンにより,遺伝子発現に大きな変化が生じるが,転写量が減少した遺伝子は増加した遺伝子より多かった39).このことからH2A.Bヌクレオソームは転写促進に機能しており,伸長を促進するRNAポリメラーゼIIとの相互作用によってより容易に破壊される安定性の低い性質が原因となっていることが考えられる.
macroH2AはN末端にmacroドメインを持つヒストンバリアントである.macroドメインはH2Bとの二量体形形成には関与しない.macroH2AはX染色体の不活性化には必要ではないが,不活性化された女性X染色体,細胞老化特異的ヘテロクロマチンに局在することから40),転写抑制に関与していると考えられる.試験管内ではmacroH2Aは転写を抑制する41).ヒトB細胞株においてIL8の転写やマウスの内在性レトロウイルスを抑制している42).macroH2Aをノックダウンすると,HOXAクラスターの遺伝子のレチノイン酸に対する感受性が上昇し,転写を抑制する作用があることが示された43).ヒト乳がん細胞株MCF-7では,macroH2Aが局在している領域の遺伝子のほとんどが発現していないが,macroH2Aを欠失させても,それらの遺伝子は発現上昇を起こさなかった.一方で,血清飢餓誘発遺伝子などの活性化にmacroH2Aが機能することも報告されている44).現在のところ,このようなmacroH2Aがどのようなメカニズムで転写の抑制と活性化の両方に機能するのかは不明である.
ヒトのゲノムDNAの解読は,遺伝子の発現制御機構の理解においてはDNA配列によらないエピジェネティクス機構の解明が必要であることを示した.エピジェネティクスはクロマチンに加えられたDNAのメチル化およびヒストンの化学修飾などの小さな変化がきわめて重要な役割を果たす.本稿で紹介したヒストンバリアントもこれらと同様にその局在がエピジェネティクスに与える影響は大きい.これらのエピゲノム情報は,単独で機能しているわけではなく,クロストークが存在している.今後の解析では,このクロストークの解明も一つの課題であると考えられる.2021年にはテロメアのDNA伸長を担うテロメラーゼのホロ酵素にH2A–H2B複合体が含まれることが発表された.これは,クロマチンに取り込まれて初めてヒストンは機能するとは限らないことを示しており,ヒストンの多岐にわたる機能を再認識させられ,これからの研究の発展が期待される.
本総説は2021年度奨励賞を受賞した.
1) Luger, K., Mäder, A.W., Richmond, R.K., Sargent, D.F., & Richmond, T.J. (1997) Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature, 389, 251–260.
2) Spiker, S., Murray, M.G., & Thompson, W.F. (1983) DNase I sensitivity of transcriptionally active genes in intact nuclei and isolated chromatin of plants. Proc. Natl. Acad. Sci. USA, 80, 815–819.
3) Passarge, E. (1979) Emil Heitz and the concept of heterochromatin: Longitudinal chromosome differentiation was recognized fifty years ago. Am. J. Hum. Genet., 31, 106–115.
4) Pombo, A. & Dillon, N. (2015) Three-dimensional genome architecture: Players and mechanisms. Nat. Rev. Mol. Cell Biol., 16, 245–257.
5) Hori, T., Amano, M., Suzuki, A., Backer, C.B., Welburn, J.P., Dong, Y., McEwen, B.F., Shang, W.H., Suzuki, E., Okawa, K., et al. (2008) CCAN makes multiple contacts with centromeric DNA to provide distinct pathways to the outer kinetochore. Cell, 135, 1039–1052.
6) Tachiwana, H., Müller, S., Blümer, J., Klare, K., Musacchio, A., & Almouzni, G. (2015) HJURP involvement in de novo CenH3(CENP-A) and CENP-C recruitment. Cell Rep., 11, 22–32.
7) Mendiburo, M.J., Padeken, J., Fülöp, S., Schepers, A., & Heun, P. (2011) Drosophila CENH3 is sufficient for centromere formation. Science, 334, 686–690.
8) Logsdon, G.A., Barrey, E.J., Bassett, E.A., DeNizio, J.E., Guo, L.Y., Panchenko, T., Dawicki-McKenna, J.M., Heun, P., & Black, B.E. (2015) Both tails and the centromere targeting domain of CENP-A are required for centromere establishment. J. Cell Biol., 208, 521–531.
9) Tachiwana, H., Kagawa, W., Shiga, T., Osakabe, A., Miya, Y., Saito, K., Hayashi-Takanaka, Y., Oda, T., Sato, M., Park, S.Y., et al. (2011) Crystal structure of the human centromeric nucleosome containing CENP-A. Nature, 476, 232–235.
10) Takizawa, Y., Ho, C.H., Tachiwana, H., Matsunami, H., Kobayashi, W., Suzuki, M., Arimura, Y., Hori, T., Fukagawa, T., Ohi, M.D., et al. (2020) Cryo-EM structures of centromeric tri-nucleosomes containing a central CENP-A nucleosome. Structre, 28, 44–53.e4.
11) Kato, H., Jiang, J., Zhou, B.R., Rozendaal, M., Feng, H., Ghirlando, R., Xiao, T.S., Straight, A.F., & Bai, Y. (2013) A conserved mechanism for centromeric nucleosome recognition by centromere protein CENP-C. Science, 340, 1110–1113.
12) Chittori, S., Hong, J., Saunders, H., Feng, H., Ghirlando, R., Kelly, A.E., Bai, Y., & Subramaniam, S. (2018) Structural mechanisms of centromeric nucleosome recognition by the kinetochore protein CENP-N. Science, 359, 339–343.
13) Foltz, D.R., Jansen, L.E., Bailey, A.O., Yates, J.R. 3rd, Bassett, E.A., Wood, S., Black, B.E., & Cleveland, D.W. (2009) Centromere-specific assembly of CENP-a nucleosomes is mediated by HJURP. Cell, 137, 472–484.
14) Dunleavy, E.M., Roche, D., Tagami, H., Lacoste, N., Ray-Gallet, D., Nakamura, Y., Daigo, Y., Nakatani, Y., & Almouzni-Pettinotti, G. (2009) HJURP is a cell-cycle-dependent maintenance and deposition factor of CENP-A at centromeres. Cell, 137, 485–497.
15) Barnhart, M.C., Kuich, P.H., Stellfox, M.E., Ward, J.A., Bassett, E.A., Black, B.E., & Foltz, D.R. (2011) HJURP is a CENP-A chromatin assembly factor sufficient to form a functional de novo kinetochore. J. Cell Biol., 194, 229–243.
16) Lacoste, N., Woolfe, A., Tachiwana, H., Garea, A.V., Barth, T., Cantaloube, S., Kurumizaka, H., Imhof, A., & Almouzni, G. (2014) Mislocalization of the centromeric histone variant CenH3/CENP-A in human cells depends on the chaperone DAXX. Mol. Cell, 53, 631–644.
17) Jeffery, D., Gatto, A., Podsypanina, K., Renaud-Pageot, C., Ponce, L.R., Bonneville, L., Dumont, M., Fachinetti, D., & Almouzni, G. (2021) CENP-A overexpression promotes distinct fates in human cells, depending on p53 status. Commun. Biol., 4, 417.
18) Ueda, J., Harada, A., Urahama, T., Machida, S., Maehara, K., Hada, M., Makino, Y., Nogami, J., Horikoshi, N., Osakabe, A., et al. (2017) Testis-specific histone variant H3t gene is essential for entry into spermatogenesis. Cell Rep., 18, 593–600.
19) Tachiwana, H., Kagawa, W., Osakabe, A., Kawaguchi, K., Shiga, T., Hayashi-Takanaka, Y., Kimura, H., & Kurumizaka, H. (2010) Structural basis of instability of the nucleosome containing a testis-specific histone variant, human H3T. Proc. Natl. Acad. Sci. USA, 107, 10454–10459.
20) Shires, A., Carpenter, M.P., & Chalkley, R. (1975) New histones found in mature mammalian testes. Proc. Natl. Acad. Sci. USA, 72, 2714–2718.
21) Montellier, E., Boussouar, F., Rousseaux, S., Zhang, K., Buchou, T., Fenaille, F., Shiota, H., Debernardi, A., Héry, P., Curtet, S., et al. (2013) Chromatin-to-nucleoprotamine transition is controlled by the histone H2B variant TH2B. Genes Dev., 27, 1680–1692.
22) Padavattan, S., Shinagawa, T., Hasegawa, K., Kumasaka, T., Ishii, S., & Kumarevel, T. (2015) Structural and functional analyses of nucleosome complexes with mouse histone variants TH2a and TH2b, involved in reprogramming. Biochem. Biophys. Res. Commun., 464, 929–935.
23) Shinagawa, T., Takagi, T., Tsukamoto, D., Tomaru, C., Huynh, L.M., Sivaraman, P., Kumarevel, T., Inoue, K., Nakato, R., Katou, Y., et al. (2014) Histone variants enriched in oocytes enhance reprogramming to induced pluripotent stem cells. Cell Stem Cell, 14, 217–227.
24) Malik, H.S. & Henikoff, S. (2003) Phylogenomics of the nucleosome. Nat. Struct. Biol., 10, 882–891.
25) Filipescu, D., Szenker, E., & Almouzni, G. (2013) Developmental roles of histone H3 variants and their chaperones. Trends Genet., 29, 630–640.
26) Ahmad, K. & Henikoff, S. (2002) The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell, 9, 1191–1200.
27) Pina, B. & Suau, P. (1987) Changes in histones H2A and H3 variant composition in differentiating and mature rat brain cortical neurons. Dev. Biol., 123, 51–58.
28) McKittrick, E., Gafken, P.R., Ahmad, K., & Henikoff, S. (2004) Histone H3.3 is enriched in covalent modifications associated with active chromatin. Proc. Natl. Acad. Sci. USA, 101, 1525–1530.
29) Tachiwana, H., Osakabe, A., Shiga, T., Miya, Y., Kimura, H., Kagawa, W., & Kurumizaka, H. (2011) Structures of human nucleosomes containing major histone H3 variants. Acta Crystallogr. D Biol. Crystallogr., 67, 578–583.
30) Mito, Y., Henikoff, J.G., & Henikoff, S. (2007) Histone replacement marks the boundaries of cis-regulatory domains. Science, 315, 1408–1411.
31) Goldberg, A.D., Banaszynski, L.A., Noh, K.M., Lewis, P.W., Elsaesser, S.J., Stadler, S., Dewell, S., Law, M., Guo, X., Li, X., et al. (2010) Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell, 140, 678–691.
32) Tagami, H., Ray-Gallet, D., Almouzni, G., & Nakatani, Y. (2004) Histone H3.1 and H3.3 complexes mediate nucleosome assembly pathways dependent or independent of DNA synthesis. Cell, 116, 51–61.
33) Drane, P., Ouararhni, K., Depaux, A., Shuaib, M., & Hamiche, A. (2010) The death-associated protein DAXX is a novel histone chaperone involved in the replication-independent deposition of H3.3. Genes Dev., 24, 1253–1265.
34) Allis, C.D., Glover, C.V., Bowen, J.K., & Gorovsky, M.A. (1980) Histone variants specific to the transcriptionally active, amitotically dividing macronucleus of the unicellular eucaryote, tetrahymena thermophila. Cell, 20, 609–617.
35) Tachiwana, H., Dacher, M., Maehara, K., Harada, A., Seto, Y., Katayama, R., Ohkawa, Y., Kimura, H., Kurumizaka, H., & Saitoh, N. (2021) Chromatin structure-dependent histone incorporation revealed by a genome-wide deposition assay. eLife, 10, e66290.
36) Obri, A., Ouararhni, K., Papin, C., Diebold, M.-L., Padmanabhan, K., Marek, M., Stoll, I., Roy, L., Reilly, P.T., Mak, T.W., et al. (2014) ANP32E is a histone chaperone that removes H2A.Z from chromatin. Nature, 505, 648–653.
37) Wang, Y., Long, H., Yu, J., Dong, L., Wassef, M., Zhuo, B., Li, X., Zhao, J., Wang, M., Liu, C., et al. (2018) Histone variants H2A.Z and H3.3 coordinately regulate PRC2-dependent H3K27me3 deposition and gene expression regulation in mES cells. BMC Biol., 16, 107.
38) Arimura, Y., Kimura, H., Oda, T., Sato, K., Osakabe, A., Tachiwana, H., Sato, Y., Kinugasa, Y., Ikura, T., Sugiyama, M., et al. (2013) Structural basis of a nucleosome containing histone H2A.B/H2A.Bbd that transiently associates with reorganized chromatin. Sci. Rep., 3, 3510.
39) Tolstorukov, M.Y., Goldman, J.A., Gilbert, C., Ogryzko, V., Kingston, R.E., & Park, P.J. (2012) Histone variant H2A.Bbd is associated with active transcription and mRNA processing in human cells. Mol. Cell, 47, 596–607.
40) Costanzi, C. & Pehrson, J.R. (1998) Histone macroH2A1 is concentrated in the inactive X chromosome of female mammals. Nature, 393, 599–601.
41) Doyen, C.M., An, W., Angelov, D., Bondarenko, V., Mietton, F., Studitsky, V.M., Hamiche, A., Roeder, R.G., Bouvet, P., & Dimitrov, S. (2006) Mechanism of polymerase II transcription repression by the histone variant macroH2A. Mol. Cell. Biol., 26, 1156–1164.
42) Agelopoulos, M. & Thanos, D. (2006) Epigenetic determination of a cell-specific gene expression program by ATF-2 and the histone variant macroH2A. EMBO J., 25, 4843–4853.
43) Buschbeck, M., Uribesalgo, I., Wibowo, I., Rue, P., Martin, D., Gutierrez, A., Morey, L., Guigo, R., Lopez-Schier, H., & DiCroce, L. (2009) The histone variant macroH2A is an epigenetic regulator of key developmental genes. Nat. Struct. Mol. Biol., 16, 1074–1079.
44) Gamble, M.J., Frizzell, K.M., Yang, C., Krishnakumar, R., & Kraus, W.L. (2010) The histone variant macroH2A1 marks repressed autosomal chromatin, but protects a subset of its target genes from silencing. Genes Dev., 24, 21–32.

公益財団法人がん研究会がん研究所研究員.博士(理学).
1981年福岡県に生る.2004年早稲田大学理工学部卒業,09年同大学院理工学研究科修了,博士(理学)取得,同大学院にて助手,助教を経て12年キュリー研究所(パリ)にて博士研究員,14年早稲田大学大学院にて講師,17年より現職.
研究テーマと抱負研究テーマは,ゲノムDNAが形成しているクロマチン構造が機能するメカニズムの解析.エピジェネティクスの解析から,転写制御を中心とした生命の基本原理および疾患のメカニズムを明らかにしたいと考えている.
ウェブサイトhttps://www.jfcr.or.jp/laboratory/department/cancer_biology/www/default.htm
趣味サッカー.

This page was created on 2022-03-15T11:57:12.006+09:00
This page was last modified on 2022-04-15T17:03:38.000+09:00
このサイトは(株)国際文献社によって運用されています。