細胞性免疫の主翼を担うメインストリームT細胞は,胸腺でのランダムなT細胞抗原受容体遺伝子(T cell antigen receptor:TCR)の遺伝子組換え後に胸腺上皮細胞上の多型性のある主要組織適合抗原複合体(major histocompatibility antigen complex:MHC)クラスI(特にこちらをクラスIaと呼ぶ)抗原/ペプチド抗原あるいはII抗原/ペプチド抗原による正・負の選択過程により生成する1).その他に,遺伝子組換えの結果作られたTCRの特異性に導かれて,主に胸腺細胞に発現するMHCクラスI分子に形状は酷似するが多型性を欠くMHCクラスI様分子(クラスIb分子と呼ぶ)と非ペプチド性小分子により選択されるT細胞も存在する.後者は反応様式が自然免疫系エフェクター細胞のように迅速であるため自然T細胞と呼ばれる2).生成した自然T細胞はセミインバリアント(α鎖が不変——インバリアント——で,β鎖がある程度固定だが多様性がある組合わせ)なTCRでMHCクラスIb/抗原を認識する.MHCクラスIb分子は多型性がない(同種ではほぼ均一,異種間であっても相同性は高い)ため,研究の結果得られた知見やリガンドによる活性化などの免疫操作法は万人に適応可能となる.自然T細胞でαβTCRタイプのものの一つにNKT細胞2)(図1)があり,クラスIbのCD1dを抗原提示分子としている.本稿ではNKT細胞の機能と疾患との関連について,マウスを用いた実験結果をもとに解説したい.
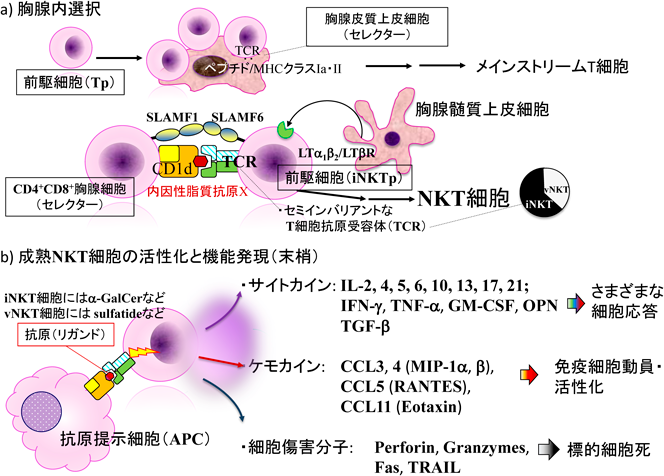
NKT細胞は,CD1d(後述)を抗原提示分子として主に脂質抗原(糖脂質,リン脂質)を認識するT細胞抗原受容体を発現する自然T細胞の一亜群である2).T細胞としてのマーカーの他にナチュラルキラー細胞のマーカーも同時に発現するところから,NKTという名称がついている3).先に述べたように,NKT細胞は自身と出自を同じくするCD4+8+(double positive:DP)胸腺細胞(骨髄由来細胞である)上のCD1d(+自己脂質抗原X)により選択され,CD1d分子に提示される脂質抗原を認識するT細胞となる4)(図1a).したがって,胸腺細胞上のCD1dを欠損(ノックアウト;KO)するとNKT細胞は生じない.通常のNKT細胞のTCRはメインストリームのT細胞と同様にα鎖とβ鎖のヘテロ二量体である(γ鎖とδ鎖を有するタイプでNKマーカー陽性のγδ NKT細胞もあるが,さらに希少なのでここでは述べない)が,このうちインバリアントα鎖を用いるNKT細胞はinvariant NKT(iNKT)細胞あるいはtype I NKT細胞と呼ばれ,NKT細胞の約6割を占める.NKT細胞のTCRの生成には,胸腺細胞におけるランダムな二度の遺伝子再構成が必要である.一度目のVβ鎖の再構成で,Vβ8, 7, 2(マウス;ヒトではVβ11など)を発現したpre T細胞が,次にVα鎖をたまたまインバリアントなVα14Jα18(マウス;ヒトではVα24Jα18)として生産的に組み換えたものがiNKT細胞として約1000個/日で生じると試算されている5).Jα18はiNKT細胞の生成に必須のJ鎖の主たるものであるので,Jα18 KOマウスはiNKT細胞を生じない6).一定の確率でVα14Jα18/Vβ7, 8, 2(invariant TCR:iTCR)+胸腺細胞(iNKT前駆細胞:iNKTp)が生じると,内因性脂質+CD1d(DP)/iTCR(iNKTp)による第1のシグナル(強いTCRシグナルである)に加え,Signaling lymphocytic activation molecule family member 1(SLAMF1)(DP)/SLAMF6(iNKTp)による第2シグナル,髄質胸腺上皮細胞(medullary thymic epithelial cell:mTEC)とのlymphotoxin β受容体(LTβR)(mTEC)/LTα1β2(iNKTp)を介し核内因子κB(nuclear factor-κB)非古典経路の活性化の下流にある第3シグナルを受けてNKT細胞の成熟が進む7–9)(図1a).この過程が通常のT細胞とは異なるエフェクター・メモリー表現型をはじめとしたメインストリームT細胞との表現型の差異を生み出すものと考えられる.CD1dで選択されるが,iTCRを発現しないNKT細胞(NKT細胞の残り4割弱)は,一部を除いてvariant TCR鎖を用いているためvNKT(あるいはtype II NKT)細胞と呼ばれる3)(図1a).
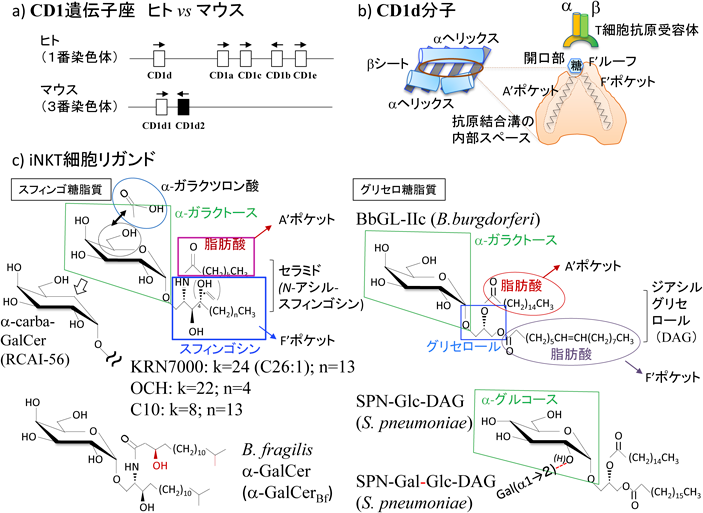
CD1dは糖脂質・リン脂質抗原の提示とNKT細胞の機能発現に重要な分子である10).MHCクラスIbの一つであるCD1分子として,ヒトはCD1a, b, cの三つのGroup I CD1(古典的CD1)とCD1d(Group II),CD1e(Group III)の二つの非古典的CD1分子を,マウスは非古典的CD1としてGroup IIのCD1dのみの遺伝子を有する(図2a)11).CD1分子はいずれも構造的に開口部は小さいが,疎水性側鎖を有するアミノ酸で形作られた深い抗原結合溝を有し,T細胞に脂質や非リボソーム性ペプチド抗原を提示する12).CD1eは細胞表面には発現せず脂質抗原提示のシャペロンとして機能する13).マウスCD1d遺伝子座にはCd1d1とCd1d2というホモロジー95%の二つの遺伝子が隣接して存在するが,さまざまな組織で発現・機能しているのは主にCD1d1である5).Cd1d2はC57BL/6(B6)マウスでは偽遺伝子となっており発現しない(α3ドメインアミノ末端に終止コドンが出現する)5).一方,BALB/c, AKR, NOD, 129などの系統ではCD1d2も少ないながら胸腺などで発現する.CD1d2はα2ドメインにシステインからトリプトファンへのアミノ酸置換変異があり14),過去のHLAクラスIa分子の同様な変異例における構造的な脆弱性から15),発現しない可能性も示唆されていた.また発現しても量や部位がCD1d1に比してかなり限定していることもあり以前はCD1d2拘束性NKT細胞の存在は想定されておらず,また実際にSv129(CD1d2発現系)/B6(CD1d2非発現系)ミックス背景でCD1d1のみをKOしてもNKT細胞の分化の大半は消失していると報告されてもいる16).しかし,in vivoでの生理機能は未知数ながら,近年詳細な免疫学的,構造生物学的な検討がなされ,CD1d2により適合する抗原やCD1d2拘束性NKT細胞[interferon(IFN)-γ産生など機能あり]の存在が確認された17).
一方,組織・臓器のCD1d1発現をNational Center for Biotechnology Information Genes and Expression Database[reads per kilobase exon per million mapped reads(RPKM)という指標がよいかは議論があるようであるが]でみてみると,皮下脂肪>肝臓>副腎>生殖腺周囲脂肪>乳腺≫胸腺>脾臓>卵巣となっており,脂肪での発現が非常に高いことがあらためてわかる18).一方,Cd1d2(非B6由来)は低い発現としてではあるが,皮下脂肪=肝臓>副腎>生殖腺周囲脂肪>乳腺>胸腺という同様な順となっている19).CD1d1(以降は特にCD1d2と区別する必要がある場合以外は単に,CD1dと記載する)とNKT細胞との相互作用については後述する.
CD1dがiNKT細胞に提示し,同細胞を活性化するプロトタイプ抗原としてα-galactosylceramide(α-GalCer)が海洋天然物agelasphinとして見いだされ20),抗腫瘍活性の根幹にあるNKT細胞の活性化リガンドであるという報告7, 21)から約4半世紀が経過した.α-GalCerは単一のリガンドでiNKT細胞からIFN-γ, interleukin(IL)-4の両方を誘導するが,その後の構造活性相関の研究からよりIFN-γ方向(α-carba-GalCer)22)あるいはIL-4の方向(OCH;KRN7000のスフィンゴシンのアルキル基長に比べ9炭素分短いリガンド)23)に偏倚した代表的なリガンドが合成され,実験に供されている(図2c).またさらなる自然抗原の探索から,ヒトやマウスの内因性糖脂質であるisoglobotriaosylceramide(iGb3)24)や腸内細菌構成成分(Bacteroides fragilis由来α-GalCer類似物質;α-GalCerBf)25, 26)にもiTCRと結合するα-Gal抗原構造が存在することが見いだされている.acylsphingosine(ceramide)の2本のアルキル鎖はCD1dのA′ (acyl)およびF′ポケット(sphingosine)に収納され,TCR面に向き合うようにα-Galが配向する27).また,グラム陰性菌でリポ多糖(lipopolysaccharide:LPS)陰性のSphingomonas属の細胞膜成分にはガラクトースの代わりにα-ガラクツロン酸が結合したセラミド28, 29)が,ライム病病原体であるらせん菌Borrelia burgdorferiではセラミドの代わりにジアシルグリセロール(DAG)をベースにα-galactosyl-DAG30)が,肺炎の重要な病原菌であるStreptococcus pneumoniaeからもSPN-Glc-DAGやSPN-Gal-Glc-DAGがiNKT細胞を刺激するリガンド31)として見いだされている.糖脂質抗原陽性の日和見感染を生じる微生物に対しては,早期のiNKT細胞による防御が重要な役割を果たすことも示されている30–32).また,現在もα-GalCerの誘導体としてiNKT細胞をさまざまに刺激し,生物活性を有する多様な化合物が合成されている33, 34).CD1d2に関してはA′ポケットが小さく,CD1d1にフィットするような長いアルキル基のα-GalCer(C26)は収まらないため,短いアシル鎖の脂肪酸のα-GalCer(C10)あるいはα-サイコシン(α-galactosylsphingosine)がよいリガンドとなることが示された17).また,vNKT細胞については神経など生体内に存在するsulfatide35)やlysosulfatide36)がCD1dにより提示され,その認識抗原となっている.いずれにせよ,図2にはそのごく一部しか掲げられないため,リガンドの化学的詳細は文献に当たられたい37, 38).
iNKT細胞全体を活性化(リガンドin vivo投与,あるいは脾細胞や肝リンパ球全体,さらにはソートしたα-GalCer負荷CD1d-tetramer+細胞をリガンドと培養)した場合,迅速・大量・多種類のサイトカイン[IL-2, 4, 5, 6, 10, 13, 17, 21, IFN-γ, tumor necrosis factor(TNF)-α, granulocyte-macrophage colony stimulating factor(GM-CSF),transforming growth factor(TGF)-β]の産生,ケモカイン[CC-chemokine ligand-3(CCL3),CCL4, CCL5, LT, eotaxin]の産生,細胞傷害装置[TNF-related apoptosis-inducing ligand(TRAIL),Fas ligand(FasL),Perforin, Granzyme]の誘導などを生じ(図1b),その多機能性から免疫細胞界のスイスアーミーナイフと表現されたこともある39).迅速な反応は活性化・メモリーマーカーを発現していることを体現する応答であるが,多種類の遺伝子発現は集団としてみた場合の平均化された全方位的な応答といえる.ハイブリドーマなどにして個々の細胞としてサイトカインなどの産生でみると,一つのクローンが産生するサイトカインにはある程度偏りが認められる40, 41).これは,均一な細胞群と想定されながらも,胸腺内選択の時点でiTCRのアビディティーなどによりNKT1/2/17に分化することで決定されているためと考えられる42).
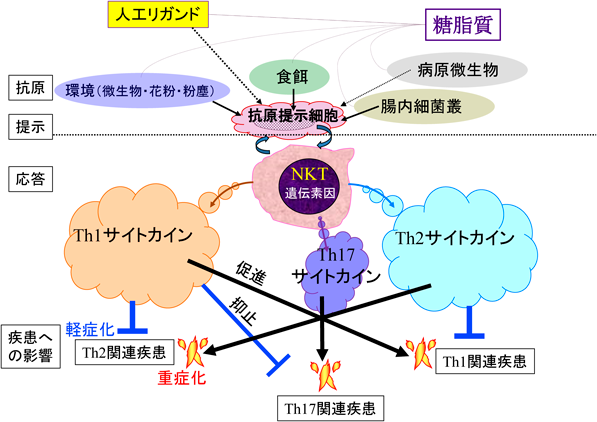
何らかの理由で自己寛容状態が破れ,そのT細胞応答がT helper 1(Th1)/Th2/Th17のいずれかへ偏倚していることが疾患の特徴であるとすれば,そのような偏倚に傾かないようにバランスをとることが寛解へとつながることがある.たとえばTh1へ,あるいはTh2へ,またTh17へ誘導される応答(それぞれ1~3型免疫偏倚)を抑止すればそれぞれの関連疾患は軽症化する(図3)43).また一見,免疫学的機序が関連しないと考えられる疾患に関しても,傷害された細胞から傷害関連分子パターン(damage-associated molecular patterns:DAMPs)44)が漏出することにより炎症性変化が惹起される.その場合も同様に1~3型免疫偏倚が進展過程を是正することは軽症化につながる.その意味で,NKT細胞の応答特性としてネガティブフィードバック制御が反応の場のサイトカイン環境とは反対の方向へ舵を切ることに役立っていると思われる45, 46).
NKT細胞(特にiNKT細胞)の活性を修飾しうる体内への摂取物質として,食餌(主に乳製品)から年間150 gくらい摂取する糖脂質(β結合のものがほとんど)47)がある.食用キノコ(そのうちのごく一部かもしれないが)にiNKT細胞を刺激する成分を有するという報告もある48).さらに,環境抗原(花粉49),土壌や水中の環境微生物50),粉塵51)),腸内細菌叢(前述のB. fragilis由来各種α-GalCer;これらが主たる供給源かもしれない)からも摂取可能である25, 26).また感染時には病原微生物などに由来する糖脂質28–31)による機能修飾(活性化,抑制),またリガンドであるα-GalCer, OCHなどによる活性化で迅速・大量・多種類のサイトカイン/ケモカイン産生や傷害活性の誘導を通して,疾患に関連する免疫応答を抑制性~促進性までさまざまに修飾しうる(図3).リガンドで人工的に刺激する以外の要因は,それぞれの個体の置かれた環境や食を中心とした生活習慣などが当たるであろう.このことを証明するように宿主の摂取した分岐アミノ酸からB. fragilisが取り込み,sphinganineを経由し,自身が合成した分岐リガンドが宿主のiNKT細胞に制御的に作用しうることが示された52).
感染症を中心にNKT細胞と多くの疾患モデルの関連については過去にかなり網羅的にまとめられ53),最近アップデートもされている54).その関連は,NKT細胞が欠損しているマウス(CD1d1遺伝子ノックアウトマウス;以下単にCD1d KO),さらにより狭くはiNKT細胞欠損(Jα18 KOマウス)で誘導した場合に疾患が重症化,あるいはα-GalCerでiNKT細胞を活性化した場合に軽症化が観察されるものはNKT細胞が疾患の軽症化に関与する(=善玉として機能),あるいはCD1d KOで軽症化するものはNKT細胞が疾患の重症化に関与する(=悪玉として機能)ということで検証されてきた.ただ,疾患モデルによっては研究室間で結果が逆となっていることがあり,注意が必要である.自験例では前者として実験的自己免疫性ぶどう膜網膜炎(experimental autoimmune uveoretinitis:EAU)55, 56),心筋梗塞後リモデリング57),心筋虚血再灌流傷害58),圧負荷心肥大・心不全59),アンギオテンシンII誘導性腹部大動脈瘤60),II型コラーゲン誘導性関節炎61),後者として食餌誘導性肥満(diet-induced obesity:DIO)62–65),concanavalin A hepatitis(自己免疫性肝炎のモデル)66, 67),動脈硬化症68, 69)などがあり,本稿ではEAUとDIOについて例示したい.
6. 実験的自己免疫性ぶどう膜網膜炎(EAU)ではiNKT細胞は善玉
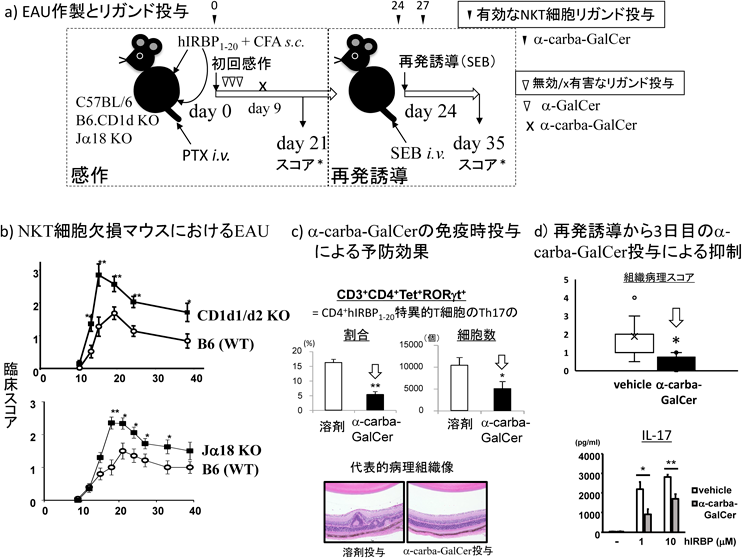
EAUはB6マウス(H-2b)をヒト視細胞間レチノイド結合タンパク質(human interphotoreceptor retinoid binding protein:hIRBP)のアミノ末端から1~20番残基ペプチド(hIRBP1–20)と完全フロイントアジュバントのエマルジョンで免疫し,同時に百日咳毒素を追加アジュバントとして投与することで誘導する(図4a左,基本型)70).通常この処置後10日前後で発症し,3週間後に眼組織病理スコアが最大となる経過でぶどう膜炎[通常ぶどう膜炎が網膜にも波及するのでぶどう膜網膜炎(uveoretinitis)ということも多い]が単相性に生じる.ベーチェット病などヒトのぶどう膜炎疾患では初発から寛解,さらに再発を繰り返すような経過をたどることが多い.そこでマウスモデルとして,ヒトのように再発を生じさせるようなモデルも考案されている71).図4a右はその一つで,ブドウ球菌エンテロトキシンB(Staphylococcal enterotoxin B:SEB)を用い,Vβ8, Vβ3などを有するhIRBP1–20特異的T細胞クローンを再活性化させ,消退しかけたぶどう膜炎を惹起させる再発誘導系である72).
EAUはCD1d KOあるいはJα18 KOマウスでは臨床スコア(経時的),組織病理スコア(免疫後21日目)とも野生型マウス[B6;wild type(WT)]より重症化した(図4b)56).これはiNKT細胞が病態進展に対して抑制的に機能していることを示唆する結果である.では,iNKT細胞活性化で軽症化が誘導されるであろうか.感作後早期にα-GalCerあるいはOCHを短回~複数回投与して検討した結果では,WTに誘導した溶剤対照群との間に臨床スコアの有意な軽症化は認められなかった.一方,感作用の乳剤中にNKT細胞リガンドを混じて皮下投与した場合,Th1方向にiNKT細胞を活性化するリガンドの投与群で21日目の組織病理スコアが溶剤対照群に比較して低下した(図4c).抗原特異的T細胞応答をhIRBP3–13/I-Adテトラマー[米国国立衛生研究所テトラマー中核施設(NIH Tetramer Core Facility:NTCF)より供与]で可視化すると,テトラマー陽性(Tet+)T細胞数には両群間に差はなかった.その反映として抗原hIRBP1–20に対する増殖反応にも両群間に有意差を認めなかった.しかし,その培養上清でIL-17, IL-22, TNF-α, IFN-γなどの産生の顕著な低下が認められた.また所属リンパ節でiNKT細胞の増加に伴ってCD4+Tet+ Th17が顕著に低下していることが示された.その結果,眼内浸潤Tet+ T細胞が21日で著明に減少した.このことは感作後早期(6時間)のリガンド投与群ではIFN-γが上昇することで所属リンパ節のTet+ Th17への分化が強く抑制されたためと考えられた.惜しむらくは,発症時付近のα-carba-GalCer投与は病態をかえって重症化させることが判明しており,有効であったのは同リガンドの予防的投与のみであった56).
そこで,再発モデルで再発誘導を行った時点とその数日後で病態制御が可能かどうかを調べた.初回感作後24日後にSEBを静脈内投与し再発を誘導すると,溶剤対照群では組織病理スコアが低下する35日で有意な増大を観察し,ぶどう膜炎の再発が認められた57).また誘導後4日でのT細胞レパトアを解析したところSEBに反応性のVβを有するT細胞が対照群と比較して増大していることが確認された(未発表データ,岩渕).同日にα-carba-GalCerを投与すると35日で溶剤投与群との間に有意差を持ってスコアの低下が認められた57).この際,初回感作と同時に投与して21日目の組織病理スコアが低下した場合と同様に所属リンパ節におけるIL-17発現が有意に低下していた.また再発誘導の3日後にα-carba-GalCerを静脈内投与した場合も組織病理スコアの著明な低下を認め,同様にIL-17産生が抑制された(図4d)57).再発誘導前にiNKT細胞に誘導されるIFN-γ産生を抗体で中和すると,IL-17産生が増強し,病理スコアは増大し,再発EAUは重症化することが示された.
初感作時に1型免疫偏倚を呈するリガンドでiNKT細胞から高レベルにIFN-γを産生させると,Th0→Th17への分化が抑制されたためIL-17産生が低下したと考えられる.再発時においてもiNKT細胞からのIFN-γ産生を作用させることで,IL-17産生抑制を伴ってEAUは軽症化した.詳細な機序の解明とリガンド投与の臨界期の決定が必要である.
DIOに取り組む契機となったのは,動脈硬化症の誘導のため当初使用していた動脈硬化食[atherogenic diet(AD);脂肪15%,コレステロール1.25%,コール酸0.5%,日本農産製]を高脂肪食[high fat diet(HFD);脂肪21.2%,コレステロール0.2%,Harlan社製)]給餌に切り替えたことによる.HFD給餌では,AD給餌で認められない体重増加を生じたが,大学院生がCD1d KOではWTやJα18 KOに比較して体重増加が鈍いことを観察した.当初,NKT細胞は緑膿菌感染防御に必要という報告もあり73)(限定的という報告も74)),免疫不全による成長不良かとも考えられた.しかし,動物施設のクリーン化後の検討でCD1d KOマウスでの体重増加の鈍化は同マウスでは脂肪組織における炎症が低く抑えられていることに起因していることが確認された.内臓脂肪における慢性炎症は肥満に重要な役割を果たしていること75, 76)は,以前よりマクロファージ(Mϕ)など自然免疫との関連で報告されてはいたが,2009年にはCD8+ T細胞77),B細胞78),調節性T細胞(regulatory T cell:Treg)79)などの獲得免疫系細胞も肥満の進展と関連があることがメジャー誌に報告されるに至って,より大きな注目を集めることとなった.肥満の進展に対してCD8+ T, B, M1 Mϕ(M1)80)は促進性に,M2 Mϕ(M2)80),Treg,好酸球81)は抑制性に機能することがこれまでに示されたわけであるが,NKT細胞に関してはいずれの可能性も考えられた.これまでの実験結果を表にまとめた(表1).CD1d欠損/NKT細胞欠損の効果が中立的なもの82),CD1d欠損の効果として悪化させるもNKT細胞非依存性のもの83),CD1d欠損が肥満,インスリン抵抗性(insulin resistance:IR)の軽快につながるNKT細胞が悪化に作用するもの(Type II NKTが悪化に関連63),Type I NKT細胞が悪化に関連84)と当初分かれていた),Vα14トランスジェニックを用いてiNKT細胞が悪化に関連するもの85),CD1d欠損が肥満,IRの悪化につながるNKT細胞が抑制的に作用するもの86–89)などが,全身性に(ゲノム上の)CD1d1あるいはCD1d1/2両方の遺伝子破壊により作られたKOの形質となっている(表1上段).これらの差がなぜ生じているかは,食餌,マウスの飼養施設の環境,腸内細菌叢(前二者の影響ともいえる),使用系統(CD1d1 KO vs CD1d/2d KO;これは現在ではほぼ否定か)などと議論されているものの,いまだ原因について確定はしていない.
表1 肥満・インスリン抵抗性に対するCD1d欠損(全身・組織特異的)の影響| 全身でのノックアウト |
|---|
| NKTの作用 | 遺伝背景 | マウス系統 | 脂肪原料 | CD1d欠損の効果 | 文献 |
|---|
| 中立的 | BALB/c | CD1d1/2 KO | 大豆油・ラード | なし | 82 |
| C57BL/6 | CD1d1/2 KO | NKT非依存性に悪化 | 83 |
| 促進的 | C57BL/6 | β2m KO, WT+α-GC | | KOで軽快・α-GCでIR悪化 | 62 |
| CD1d1 KO | ベニ花油・牛脂 | 肥満・IRは軽快 | 63, 91 |
| CD1d1 KO | 大豆油・ラード | 肥満・IRは軽快 | 84 |
| Vα14Tg/LDLR KO | ラード・ショ糖・コレステロール(0.15%) | Tgで肥満・IRは悪化 | 85 |
| 抑制的 | C57BL/6 | CD1d1/2 KO | 大豆油・ラード | 肥満・IRは悪化 | 86 |
| CD1d1/2 KO | 肥満・IRは悪化 | 87 |
| CD1d1/2 KO | 肥満・IRは悪化 | 88 |
| CD1d1/2 KO | 肥満・IRは悪化 | 89 |
| コンディショナルノックアウト(すべてC57BL/6背景) |
|---|
| NKTの作用 | 欠損細胞 | creの駆動遺伝子 | 脂肪原料 | CD1d欠損の効果 | 文献 |
|---|
| 促進的 | 脂肪細胞 | Adipoq-cre | ベニ花油・牛脂 | 肥満・IRは改善 | 64 |
| 抑制的 | 大豆油・ラード | 肥満・IRは悪化 | 103 |
| 促進的 | マクロファージ | LysM-cre | ベニ花油・牛脂 | 肥満・IRは改善 | 65 |
| 大豆油・ラード* | 肥満・IRは改善 | 101 |
| 抑制的 | 樹状細胞? | CD11c-cre | ベニ花油・牛脂 | 肥満・IRは悪化 | 65 |
| M1 Mϕ | 大豆油・ラード* | 肥満・IRは悪化 | 101 |
| これまでの,主としてCD1d1あるいはCD1d1/d2のゲノムKOおよびCD1d1fl/flと組織・細胞系譜特異的発現creとの交配によるコンディショナルKOの欠損の効果を表にした.マウスの遺伝的背景と系統,餌の脂質の原料,CD1d欠損等の一次的効果,文献番号を記した.脂質の由来不明は空欄となっている.*は文献の記載から推定した. |
8. 食餌誘導性肥満(DIO)ではNKT細胞は悪玉か?
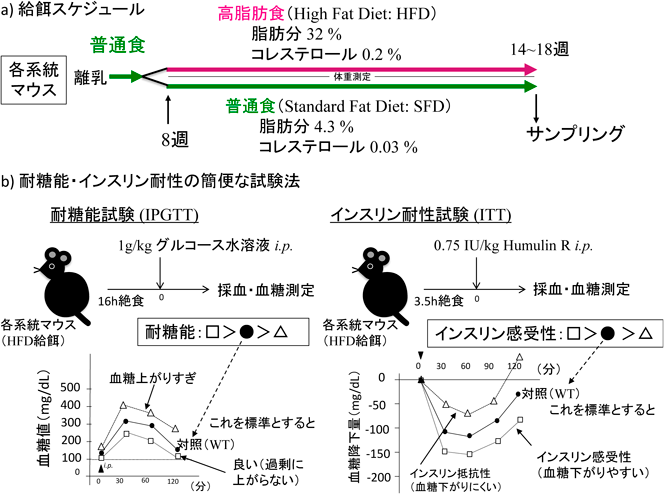
我々はHFD[日本クレア製HFD32;脂肪32%(牛脂15.88%,ベニバナ油20%),全体5.08 kcal/g]を図5aのスケジュールでマウスに給餌した63).また,耐糖能は腹腔内投与グルコース耐性試験(intraperitoneal glucose tolerance test:IPGTT)で,インスリン感受性(抵抗性)は高インスリン正常血糖クランプ法などの精密な試験ではないが,インスリン耐性試験(insulin tolerance test:ITT)で評価した(図5b).
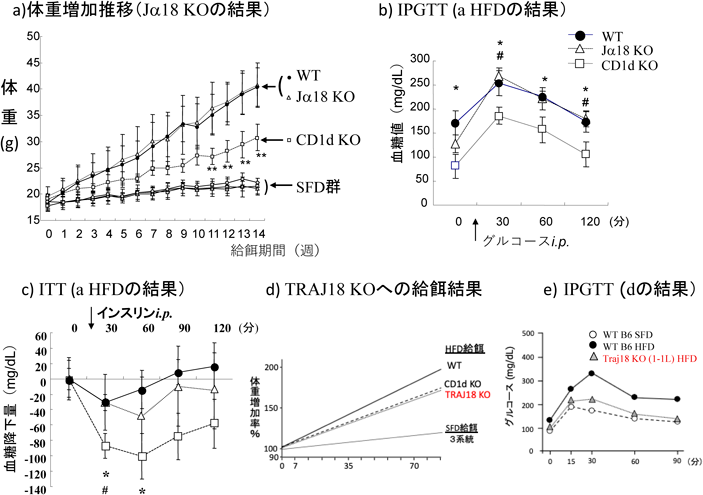
B6(WT),Jα18 KO(iNKT細胞欠損),CD1d1 KO(全NKT細胞欠損)の3系統のマウスについて調べた結果,WTとJα18KOは同等の体重増加を示し,その内訳たる生殖腺周囲脂肪重量,肝重量もWTと同等で,対するCD1d KOの場合よりも有意な増加を呈した(図6a)63).脂肪細胞周囲径(顕微計測),肝脂肪蓄積量(オイルレッドO染色)もWT=Jα18 KO>CD1d KOであった.肥満を反映して,インスリン感受性が低下し,グルコースの負荷後WTとJα18 KOは同レベルの高血糖が認められたのに対して,CD1d KOのIPGTTは非常に低く抑えられ(図6b),インスリン投与後の血糖降下も著明であった(図6c).また,WT, Jα18 KOの脂肪組織のTCRβ+NK1.1+細胞はHFD給餌で同様の割合で増加しており,Jα18 KOではCD8+亜群がWTやCD1d KOに比較して割合が増加していた.またVβ使用ではVβ5がWT, Jα18 KOで比較的多く認められた.これらの特徴から応答NKT細胞亜群は明らかにしてはいなかったもののiNKT細胞がなくともvNKTが脂肪織炎症を惹起し,肥満の進展を支持・促進していると報告した63).また,Jα18 KO肝単核球をCD1d KOに移入したものでは,脂肪にCD8+TCRβ+NK1.1+細胞が有意に出現したこと,体重も有意差を生じるほどでないがCD1d KOマウス由来肝単核球をCD1d KOに移入した対照群を下回ることはなかったこと,その反映としてITT試験で抵抗性が移入されたことなど,Jα18 KOに脂肪織炎症を惹起するvNKT細胞が存在する可能性も併せて示した63).しかし,その後Jα18 KO(Jα281 KO)は挿入遺伝子の影響でかなりの数のJαが再構成に使用されないことが報告された90)(研究室のJα18 KOでも調べたところ当然ながら同様であった).そこで新たにCRISPR/Cas9でJα18のみをgRNAで破壊したTRAJ18 KOの1系統の供与を得て,DIOを誘導91)したところ,このiNKT細胞欠損マウスでは,CD1d KOと同様の体重増加,IPGTT(図6d, e)を示した.つまり,肥満の進展を支持する十分な炎症を惹起するのはiNKT細胞であるというWuらの結果と合致した85).先のJα18 KOでWT並みに肥満が進展したのかメカニズムは不明のままであり,肥満・インスリン抵抗性へのvNKT細胞の関与は,移入に用いたHFD給餌Jα18 KO肝リンパ球および内臓脂肪NKT細胞のレパトアの解析を待たねばならない.
9. 食餌誘導性肥満(DIO)ではNKT細胞は善玉か?
一方,DIOをWTとCD1d KOあるいはJα18 KOマウスに誘導した場合,かえって肥満,IRが増悪・進展したと報告したグループは,NKT細胞がIL-10を産生し脂肪織炎症を抑制,インスリン感受性は保たれると報告している87).また,脂肪組織に局在するNKT細胞(adipose tissue NKT:AT-NKT)は,他の組織のNKT細胞がpromyelocytic leukemia zinc finger(PLZF)を特徴的な転写因子として分化するのに対し,transcriptional repressor E4-binding protein 4[E4BP4;nuclear factor, IL-3-regulated(NF-IL3)と同一]が用いられるとしている92).E4BP4(NF-IL3)は,脂肪組織のサイトカイン環境をIL-10, IL-13産生性に,抗炎症性に保つ機能を発現する可能性があり,AT-NKT細胞は分化時点ですでに異なっていることを示すのかもしれない.
さらに,脂肪組織ではNKT細胞が抗炎症性サイトカイン産生により炎症を抑制するだけではなく,線維芽細胞増殖因子FGF-21を介し,白色脂肪を褐色脂肪化させ燃焼(体熱産生)し,減量するというメカニズムも示している93).AT-NKT細胞についてもNK1.1+とNK1.1−の亜群で機能の違い[(+):病的Mϕのトリミング,(−):IL-10産生]を明らかにしており94),Lynch博士らはこの分野を独走している.その他のグループはまた,微妙な点で異なりもするが,iNKT細胞が肥満,IRの誘導に対して抵抗するのは同様である86, 88, 89).
齟齬のある結果はいかんともしがたいが,彼らがマウスに給餌している高脂肪食は表1にあるように大豆油とラードからなるResearch Diets社のD12492(青い外見)の製品である.カロリーは5.24 kcal/gと同程度ながら,脂質の原料がラード31.6%と大豆油3.2%と我々が給餌したHFD32と脂肪の内容がかなり異なっている.ラードと牛脂では構成脂肪酸の組成も,前者ではC18:1, C18:2などの不飽和脂肪酸が多く,後者ではC18:0, C14:0など飽和脂肪酸が多い特徴がある95).D12492を給餌した結果,当研究室でも体重がWTを超えることはなかったが,耐糖能で見た場合KOがWTより悪化したので,給餌したHFDの違いも一つの要因かと考えている(未発表データ).しかし,WuらはそのHFDを給餌してiNKT細胞は促進的と報告している84).
HFDの他に密接に関連し,かつ容易にコントロールできない腸内細菌叢の問題がある.SFD給餌ではWT, CD1d KOともに腸内細菌叢がBacteroidetes門細菌>Firmicutes門細菌であるが,HFDの給餌期間が永くなることにより次第にFirmicutes門細菌>Bacteroidetes門細菌となる.この置換がCD1d KOでは遅く,また属レベルでみるとより違いが認められた(未発表データ).CD1d-NKT細胞軸が腸内細菌の定着に関連するという報告96)から考えると,細菌叢の違いはCD1d KOにおける肥満,IRとも関連しているのではないかと思われるが,証明には無菌化マウスへの注目する菌の単一移植系を構築するなどの必要があり,課題となっている.
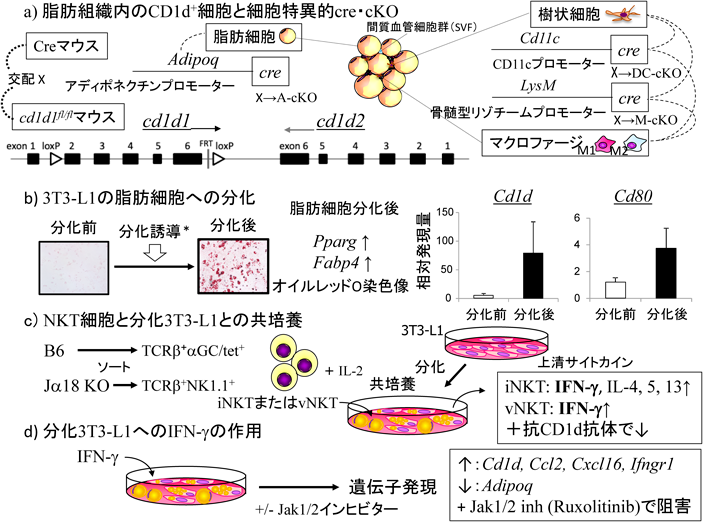
我々の結果からは,全身的CD1d KOでDIOの進展が軽減されることが判明し,特にiNKT細胞が何らかの内因性リガンドを認識・活性化することで,脂肪組織における慢性炎症の基盤を形成していることが推測された.内因性リガンドの解明が最も重要と考えられるが,まず脂肪組織中の主たる抗原提示細胞は何かを明らかにする実験に着手した.脂肪組織中で主にCD1dを発現する細胞としては,脂肪細胞,次いで大きな脂肪細胞がひしめいている隙間に間質血管細胞群(stromal vascular fraction:SVF)として存在するマクロファージ,樹状細胞などがある.CD1d1 floxed mouse(cd1d1fl/fl)はBaiらによりすでに作製されており97),また細胞・組織特異的プロモーターで駆動されるCreマウスも幾種類も樹立されていたので,これらを交配して細胞特異的CD1dコンディショナルKO(cKO)マウスを作製し,DIOを誘導・解析することとした(図7a).脂肪細胞特異的CD1d cKO(AdipoΔCD1d1; A-cKO)マウスはアディポネクチン(Adipoq)-cre98)×cd1d1fl/flマウスから,マクロファージ特異的CD1d cKO(MϕΔCD1d1; M-cKO)は骨髄型リゾチーム(myeloid lysozyme;LysM)-cre99)×cd1d1fl/flマウスから,樹状細胞特異的CD1d cKO(DCΔCD1d1; DC-cKO)マウスはCD11c-cre100)×cd1d1fl/flマウスから準備し,いずれも対照としてはfl/fl, cre(-)マウスでWT形質を示す個体を用いた.DC-cKOではCD11cを活性化マーカーと考え,樹状細胞でのCD1d KOではなく,むしろM1マクロファージのCD1d KOと捉えている見方もある101).
11. 脂肪細胞でのCD1d KO(A-cKO)のDIO
脂肪細胞はCD1dを発現するが,これは線維芽細胞3T3の中で脂肪細胞分化能を持つ3T3-L1も同様である.分化培養に従ってその発現が誘導され,同時にCD80も発現し,抗原提示能が十分に備わっていることを示す(図7b).WTからソートしたiNKT細胞は,分化3T3-L1脂肪細胞と共培養すると外からリガンドを加えなくともIFN-γ, IL-4, 5, 13を産生する64)(図7c).この産生は抗CD1d抗体で阻害されるので,脂肪細胞は何らかの内因性(脂質)抗原を発現しているものと考えられる(ただしα-GalCer/CD1d複合体をiTCRのように認識するL363抗体では検出できない).Jα18 KOからソートしたvNKT細胞もIFN-γ産生を示す.産生されたIFN-γが脂肪細胞側の遺伝子発現にどのような影響を与えるかをin vitroで調べた(図7d).マクロファージをリクルートするCcl2,NKT細胞をリクルートするCxcl16の他に,抗原提示分子CD1dやIFN-γ受容体Ifngr1などの発現が増強され,炎症がポジティブフィードバックで進展する可能性が示された.
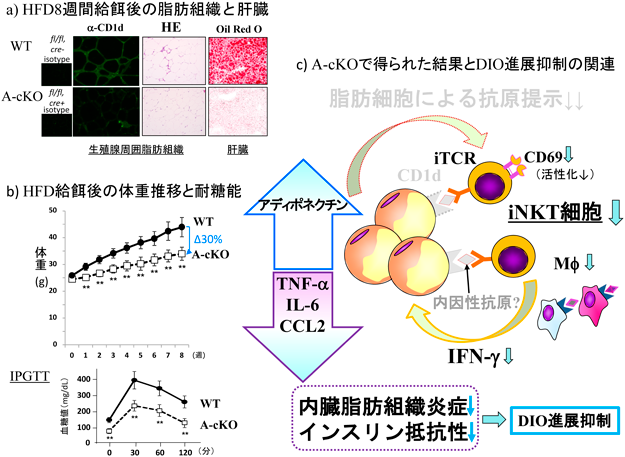
A-cKO(図7a)が樹立されると,胸腺細胞でのCD1d発現は正常であるので,NKT細胞は分化し,それぞれの局在部位に配置されているが,HFD給餌しても活性化(CD69+)NKT細胞,細胞内IFN-γ+ NKT細胞の割合・実数が著明に減少した64).また,内臓脂肪に分布するマクロファージ,iNKT/vNKT細胞も著明に減少し,炎症性サイトカイン・ケモカインの発現も大きく減じていた.この脂肪織炎症の「鎮火」により,脂肪細胞の肥大や肝細胞への脂質蓄積は抑制され(図8a),給餌8週後の体重増もWTより約30%減じ(図8b),これは全身でのCD1d KOと同等であった.このことは,皮下脂肪や血管周囲脂肪組織102)など身体中にあまねく分布する細胞でCD1dが除去された効果を意味しているのかもしれない.また,このA-cKOマウスとまったく同じように作製された脂肪細胞特異的CD1d1 KOマウスが別の研究室から報告されたが,結果はやはり,というべきか,逆であった(表1)103).我々のA-cKOにおける結果と脂肪組織内での細胞動態などをまとめた(図8c).
12. マクロファージでのCD1d KO(M-cKO)
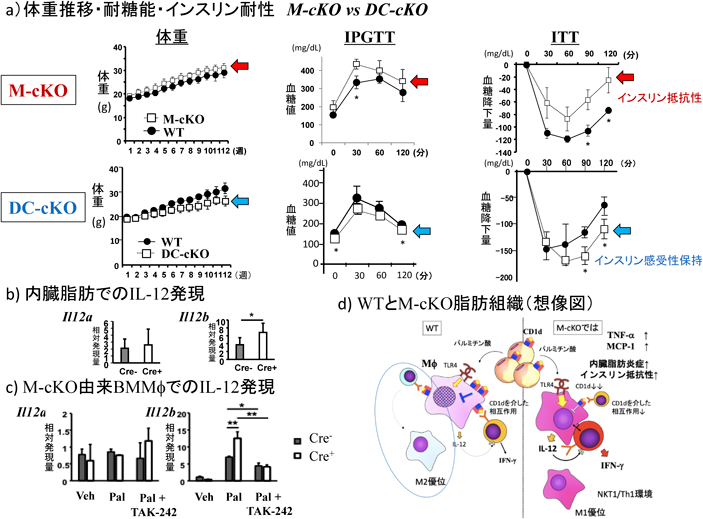
Mϕおよび樹状細胞(dendritic cell:DC)は同じ骨髄球系であることもあり,LysM-creあるいはCD11-creで,CD1d発現が除去される集団は一部重複し,またLysM-creではMϕでありながら,完全にはCD1d発現が除去できず,一部低下にとどまる集団もあった.ただし,M-cKO由来骨髄細胞から誘導したMϕ(bone marrow-derived Mϕ:BMMϕ)を抗原提示細胞としてα-GalCerとiNKT細胞と培養した場合には,TNF-α, IFN-γ, IL-4などの産生は著明に減少し,CD1d発現量は機能的には十分低下していることを確認した.また,M-cKOとDC-cKOではCD1d除去範囲が一部重複しても結果は対照的であった(図9a).結論的にいえばM-cKOでは内臓脂肪の環境はNKT1/Th1/M1の1型偏倚となり,体重は有意にWTより重いということはなかったが,下回ることはなく,脂肪細胞のサイズは有意に大きく,肝臓への脂肪蓄積もM-cKO>WTであり,耐糖能,IRはM-cKOで悪化していた65).一方,DC-cKOではM-cKOとは逆に体重はWTより低く推移し,耐糖能,IRはDC-cKOでWTより良好であった65).M-cKOでは内臓脂肪でのMϕ細胞(CD11b+F4/80hi)の割合・実数とも対照群より有意に多かった.M1/M2サブセットでみると,M1亜群が多いわけではなかったが,M2亜群の割合が減少し,比ではM1優位となっていた.これらの結果は先行グループのものと一致していた101).
この1型免疫偏倚を示す目的で,CD4+ T細胞でT-bet+細胞の優勢,さらにCD4+ T細胞の細胞内サイトカインを調べたところ,肝・内臓脂肪でIFN-γ+,IFN-γ+/TNF-α+細胞が増加していた.また,iNKT細胞をRORγtとT-betで展開してT-bet+ RORγt−(NKT1),T-bet− RORγt−(NKT2)の画分を調べると,内臓脂肪でNKT1の増加/NKT2の減少が,また細胞内IFN-γ+ iNKT細胞が増え,ソートiNKT細胞を刺激したときにIFN-γ, TNF-α産生が認められた65).この偏倚をもたらすものは何かを探る目的で,HFD給餌M-cKOのBMMϕのIL-12の構成サブユニットの遺伝子発現を調べたところIl12b(p40)の発現が増強していた(図9b).in vivoでのMϕを刺激する非病原微生物由来TLRリガンドとして作用するものとしてパルミチン酸を想定し,M-cKOのBMMϕを刺激するとIl12bの発現増強が観察された.この増強は,TLR4アンタゴニストTAK-242で阻害された(図9c).M-cKO BMMϕとiNKT細胞をパルミチン酸(+)で共培養すると,細胞間接触のある状態で培養した方が,Il12bの産生が増強した.また培養後非付着細胞のIFN-γ産生を調べると対照に比して増強が認められた.これらの結果をシェーマにまとめた(図9d)が,Mϕ-iNKT細胞間の相互作用を仲介する分子など不明な点も多く,ここでも今後の詳細な検討が必要である.
13. NKT細胞の関与ではなくCD1dからのネガティブシグナリングが大切?
これまでNKT細胞自体の欠損や特定の組織や細胞におけるCD1d発現の除去により,その細胞とNKT細胞の相互作用がなくなることが,実験的自己免疫性炎症モデルやHFDの給餌によるDIOやIRにどのような影響を与えるか,を考察してきた.しかし,CD1d+抗原提示細胞-NKT細胞相互作用を考えずとも,逆に特定の細胞に発現するCD1d自体が,特に炎症時などに影響を与える可能性を示唆する論文が2020年に発表された104).すなわち,同じLysM-cre x cd1d1fl/flを用いCD1dがマクロファージから除去されると,ドデシル硫酸ナトリウム(DSS)で誘導される腸炎に対してM-cKOが抵抗性になるという内容である.CD1dからの内在性シグナルが入らなくなりマクロファージへの炎症惹起時にインフラマソームの発現に影響を与えることでDSS腸炎が軽症化するというのである.この際に,CD1dに結合性のiNKT細胞リガンドでもあるiGb3の誘導やCD1dへの結合もこのシグナル経路に関連し,それでいてNKT細胞の関与はないという結果であり謎である.ただ,M-cKOマクロファージに関するいくつかの結果については,マクロファージのCD1dからのネガティブシグナリングの欠損によって説明がつく可能性もあり,この経路についてもさらなる検討が必要と考えられた.
二つのマウス疾患モデルにおけるNKT細胞の役割(あるいはCD1d分子それ自身かもしれない可能性)について紹介した.報告した研究室によらず同じ結果となる疾患(たとえばNKT細胞が促進的な役割の動脈硬化症)については,NKT細胞機能の抑制などが治療法の一つの候補となるかもしれない.しかし,研究者により帰結が逆となる結果は,それぞれの研究室での「正しい結果」と思われるが,その違いを生み出す原因がどこにあるかを明らかにしなければならない.さらにはJα18 KOとTRAJ18 KOの違いで生じた自研究室からの齟齬の究明も責務であり,この研究も回り道的ながらNKT細胞の機能発現の理解には必須であると考えている.
いずれにしても,がんについてはすでに先行しているように105),他の疾患でもいつかNKT細胞治療やリガンド・アンタゴニスト投与などが疾患治療の候補に上ることを願いたい.
これまで研究の遂行をご指導・ご協力下さいました,皆様(恩師から共同研究者,所属機関の教員・大学院生・学部卒業研究生などたくさんの方々がいらして書き切れません)にこの場をお借りして感謝申し上げます.
引用文献References
1) Klein, L., Kyewski, B., Allen, P.M., & Hogquist, K.A. (2014) Positive and negative selection of the T cell repertoire: what thymocytes see (and don’t see). Nat. Rev. Immunol., 14, 377–391.
2) Jameson, J., Witherden, D., & Havran, W.L. (2003) T-cell effector mechanisms: γδ and CD1d-restricted subsets. Curr. Opin. Immunol., 15, 349–353.
3) Godfrey, D.I., MacDonald, H.R., Kronenberg, M., Smyth, M.J., & Van Kaer, L. (2004) NKT cells: what’s in a name? Nat. Rev. Immunol., 4, 231–237.
4) Bendelac, A. (1995) Positive selection of mouse NK1+ T cells by CD1-expressing cortical thymocytes. J. Exp. Med., 182, 2091–2096.
5) Bendelac, A., Rivera, M.N., Park, S.-H., & Roark, J.H. (1997) Mouse CD1-specific NK1 T cells: Development, specificity, and function. Annu. Rev. Immunol., 15, 535–562.
6) Cui, J., Shin, T., Kawano, T., Sato, H., Kondo, E., Toura, I., Kaneko, Y., Koseki, H., Kanno, M., & Taniguchi, M. (1997) Requirement for Vα14 NKT cells in IL-12-mediated rejection of tumors. Science, 278, 1623–1626.
7) Salio, M., Silk, J.D., Jones, E.Y., & Cerundolo, V. (2014) Biology of CD1- and MR1-restricted T cells. Annu. Rev. Immunol., 32, 323–366.
8) Li, W., Sofi, M.H., Rietdijk, S., Wang, N., Terhorst, C., & Chang, C.-H. (2007) The SLAM-associated protein signaling pathway is required for development of CD4+ T cells selected by homotypic thymocyte interaction. Immunity, 27, 763–774.
9) White, A.J., Lucas, B., Jenkinson, W.E., & Anderson, G. (2018) Invariant NKT cells and control of the thymus medulla. J. Immunol., 200, 3333–3339.
10) McEwen-Smith, R.M., Salio, M., & Cerundolo, V. (2015) CD1d-dependent endogenous and exogenous lipid antigen presentation. Curr. Opin. Immunol., 34, 116–125.
11) Porcelli, S.A. (1995) The CD1 family: a third lineage of antigen-presenting molecules. Adv. Immunol., 59, 1–98.
12) Zajonc, D.M. (2014) CD1c caves in on lipids. Nat. Immunol., 19, 322–324.
13) De la Salle, H., Mariotti, S., Angenieux, C., Gilleron, M., Garcia-Alles, L.-F., Malm, D., Berg, T., Paoletti, S., Maître, B., Mourney, L., et al. (2005) Assistance of microbial glycolipid antigen processing by CD1e. Science, 310, 1321–1324.
14) Bradbury, A., Belt, K.T., Neri, T.M., Milstein, C., & Calabi, F. (1988) Mouse CD1 is distinct from and co-exists with TL in the same thymus. EMBO J., 7, 3081–3086.
15) Warburton, R.J., Matsui, M., Rowland-Jones, S.L., Gammon, M.C., Katzenstein, G.E., Wei, T., Edidin, M., Zweerink, H.J., McMichael, A.J., & Frelinger, J.A. (1994) Mutation of the alpha 2 domain disulfide bridge of the class I molecule HLA-A*0201. Effect on maturation and peptide presentation. Hum. Immunol., 39, 261–271.
16) Chen, Y.-H., Wang, B., Chun, T., Zhao, L., Cardell, S., Behar, S.M., Brenner, M.B., & Wang, C.-R. (1999) Expression of CD1d2 on thymocytes is not sufficient for the development of NK T cells in CD1d1-deficient mice. J. Immunol., 162, 4560–4566.
17) Sundararaj, S., Zhang, J., Krovi, S.H., Bedel, R., Tuttle, K.D., Veerapen, N., Besra, G.S., Khandokar, Y., Praveena, T., Le Nours, J., et al. (2018) Differing roles of CD1d2 and CD1d1 proteins in type I natural killer T cell development and function. Proc. Natl. Acad. Sci. USA, 115, E1204–E1213.
18) https://www.ncbi.nlm.nih.gov/gene/12479
19) https://www.ncbi.nlm.nih.gov/gene/12480
20) Natori, T., Akimoto, K., Motoki, K., Koezuka, Y., & Higa, T. (1997) Development of KRN7000, derived from agelasphin produced by Okinawan sponge. Nippon Yakurigaku Zasshi, 110(Suppl 1), 63–68.
21) Kawano, T., Cui, J., Koezuka, Y., Toura, I., Kaneko, Y., Motoki, K., Ueno, H., Nakagawa, R., Sato, H., Kondo, E., et al. (1997) CD1d-restricted and TCR-mediated activation of Vα14 NKT cells by glycosylceramides. Science, 278, 1626–1629.
22) Tashiro, T., Sekine-Kondo, E., Shigeura, T., Nakagawa, R., Inoue, S., Omori-Miyake, M., Chiba, T., Hongo, N., Fujii, S., Shimizu, K., et al. (2010) Induction of Th1-biased cytokine production by α-carba-GalCer, a neoglycolipid ligand for NKT cells. Int. Immunol., 22, 319–328.
23) Miyamoto, K., Miyake, S., & Yamamura, T. (2001) A synthetic glycolipid prevents autoimmune encephalomyelitis by inducing TH2 bias of natural killer T cells. Nature, 413, 531–534.
24) Zhou, D., Mattner, J., Cantu, C. 3rd, Schrantz, N., Yin, N., Gao, Y., Sagiv, Y., Hudspeth, K., Wu, Y.P., Yamashita, T., et al. (2004) Lysosomal glycosphingolipid recognition by NKT cells. Science, 306, 1786–1789.
25) Wieland-Brown, L.C., Penaranda, C., Kashyap, P.C., Williams, B.B., Clardy, J., Kronenberg, M., Sonnenburg, J.L., Comstock, L.E., Bluestone, J.A., & Fischbach, M.A. (2013) Production of α-galactosylceramide by a prominent member of human gut microbiota. PLoS Biol., 11, e1001610.
26) An, D., Oh, S.F., Olszak, T., Neves, J.F., Avci, F.Y., Erturk-Hasdemir, D., Lu, X., Zeissig, S., Blumberg, R.S., & Kasper, D.L. (2014) Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell, 156, 123–133.
27) Borg, N.A., Wun, K.S., Kjer-Nielsen, L., Wilce, M.C., Pellicci, D.G., Koh, R., Besra, G.S., Bharadwaj, M., Godfrey, D.I., McCluskey, J., et al. (2007) CD1d-lipid-antigen recognition by the semi-invariant NKT T-cell receptor. Nature, 448, 44–49.
28) Kinjo, Y., Wu, D., Kim, G., Xing, G.W., Poles, M.A., Ho, D.D., Tsuji, M., Kawahara, K., Wong, C.H., & Kronenberg, M. (2005) Recognition of bacterial glycosphingolipids by natural killer T cells. Nature, 434, 520–525.
29) Mattner, J., Debord, K.L., Ismail, N., Goff, R.D., Cantu, C. 3rd, Zhou, D., Saint-Mezard, P., Wang, V., Gao, Y., Yin, N., et al. (2005) Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature, 434, 525–529.
30) Kinjo, Y., Tupin, E., Wu, D., Fujio, M., Garcia-Navarro, R., Benhnia, M.R., Zajonc, D.M., Ben-Menachem, G., Ainge, G.D., Painter, G.F., et al. (2006) Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat. Immunol., 7, 978–986.
31) Kinjo, Y., Illarionov, P., Vela, J.L., Pei, B., Girardi, E., Li, X., Li, Y., Imamura, M., Kaneko, Y., Okawara, A., et al. (2011) Invariant natural killer T cells recognize glycolipids from pathogenic Gram-positive bacteria. Nat. Immunol., 12, 966–974.
32) Behar, S.M. & Porcelli, S.A. (2007) CD1-restricted T cells in host defense to infectious diseases. Curr. Top. Microbiol. Immunol., 314, 215–250.
33) Inuki, S., Hirata, N., Kashiwabara, E., Kishi, J., Aiba, T., Teratani, T., Nakamura, W., Kojima, Y., Maruyama, T., Kanai, T., et al. (2020) Polar functional group-containing glycolipid CD1d ligands modulate cytokine-biasing responses and prevent experimental colitis. Sci. Rep., 10, 15766.
34) Saavedra-Avila, N.A., Keshipeddy, S., Guberman-Pfeffer, M.J., Pérez-Gallegos, A., Saini, N.K., Schäfer, C., Carreño, L.J., Gascón, J.A., Porcelli, S.A., & Howell, A.R. (2020) Amide-Linked C4″-saccharide modification of KRN7000 provides potent stimulation of human invariant NKT cells and anti-tumor immunity in a humanized mouse model. ACS Chem. Biol., 15, 3176–3186.
35) Zajonc, D.M., Maricic, I., Wu, D., Halder, R., Roy, K., Wong, C.H., Kumar, V., & Wilson, I.A. (2005) Structural basis for CD1d presentation of a sulfatide derived from myelin and its implications for autoimmunity. J. Exp. Med., 202, 1517–1526.
36) Roy, K.C., Maricic, I., Khurana, A., Smith, T.R.F., Halder, R.C., & Kumar, V. (2008) Involvement of secretory and endosomal compartments in presentation of an exogenous self-glycolipid to type II NKT cells. J. Immunol., 180, 2942–2950.
37) Brutkiewicz, R.R. (2006) CD1d ligands: The good, the bad, and the ugly. J. Immunol., 177, 769–775.
38) Tashiro, T. (2012) Structure–activity relationship studies of novel glycosphingolipids that stimulate natural killer T-cells. Biosci. Biotechnol. Biochem., 76, 1055–1067.
39) Matsuda, J.L., Mallevaey, T., Scott-Browne, J., & Gapin, L. (2008) CD1d-restricted iNKT cells, the ‘Swiss-Army knife’ of the immune system. Curr. Opin. Immunol., 20, 358–368.
40) Nyambayar, D., Iwabuchi, K., Hedlund, E., Murakawa, S., Shirai, K., Iwabuchi, C., Kon, Y., Miyazaki, Y., Yanagawa, Y., & Onoé, K. (2007) Characterization of NKT-cell hybridomas expressing invariant T-cell antigen receptors. J. Clin. Exp. Hematop., 47, 1–8.
41) Satoh, M., Saeki, M., Yoshino, K., Mita, K., Iizuka, M., Hattori, A., Takeuchi, E., Eshima, K., & Iwabuchi, K. (2020) De novo generation of CD1d1-deficient NKT-cell hybridomas from CD1d−/− mice or by gene editing of CD1+ hybridomas. Kitasato Med. J., 50, 34–43.
42) Hogquist, K. & Georgiev, H. (2020) Recent advances in iNKT cell development. F1000 Res., 9, F1000 Faculty Rev-127.
43) Lim, J.Y., Park, M.J., Im, K.I., Kim, N., Jeon, E.J., Kim, E.J., Cho, M.L., & Cho, S.G. (2014) Combination cell therapy using mesenchymal stem cells and regulatory T-cells provides a synergistic immunomodulatory effect associated with reciprocal regulation of Th1/ Th2 and Th17/Treg cells in a murine acute graft-versus-host disease model. Cell Transplant., 23, 703–714.
44) Zindel, J. & Kubes, P. (2020) DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol., 15, 493–518.
45) Ota, T., Takeda, K., Akiba, H., Hayakawa, Y., Ogasawara, K., Ikarashi, Y., Miyake, S., Wakasugi, H., Yamamura, T., Kronenberg, M., et al. (2005) IFN-γ mediated negative feedback regulation of NKT-cell function by CD94/NKG2. Blood, 106, 184–192.
46) Minami, K., Yanagawa, Y., Iwabuchi, K., Shinohara, N., Harabayashi, T., Nonomura, K., & Onoé, K. (2005) Negative feedback regulation of T helper type 1 (Th1)/Th2 cytokine balance via dendritic cell and natural killer T cell interactions. Blood, 106, 1685–1693.
47) Vesper, H., Schmelz, E.M., Nikolova-Karakashian, M.N., Dillehay, D.L., Lynch, D.V., & Merrill, A.H. Jr. (1999) Sphingolipids in food and the emerging importance of sphingolipids to nutrition. J. Nutr., 129, 1239–1250.
48) Nozaki, H., Itonori, S., Sugita, M., Nakamura, K., Ohba, K., Ohnishi, M., Imai, K., Igarashi, Y., Suzuki, A., & Kushi, Y. (2010) Invariant Vα14 natural killer T cell activation by edible mushroom acidic glycosphingolipids. Biol. Pharm. Bull., 33, 580–584.
49) Dahl, Å. (2018) Pollen lipids can play a role in allergic airway inflammation. Front. Immunol., 9, 2816.
50) Kawahara, K., Kuraishi, H., & Zähringer, U. (1999) Chemical structure and function of glycosphingolipids of Sphingomonas spp and their distribution among members of the α-4 subclass of Proteobacteria. J. Ind. Microbiol. Biotechnol., 23, 408–413.
51) Finkelman, F.D., Yang, M., Orekhova, T., Clyne, E., Bernstein, J., Whitekus, M., Diaz-Sanchez, D., & Morris, S.C. (2004) Diesel exhaust particles suppress in vivo IFN-gamma production by inhibiting cytokine effects on NK and NKT cells. J. Immunol., 172, 3808–3813.
52) Oh, S.F., Praveena, T., Song, H., Yoo, J.S., Jung, D.J., Erturk-Hasdemir, D., Hwang, Y.S., Lee, C.C., Le Nours, J., Kim, H., et al. (2021) Host immunomodulatory lipids created by symbionts from dietary amino acids. Nature, 600, 302–307.
53) Yu, K.O.A. & Porcelli, S.A. (2005) The diverse function of CD1d-restricted NKT cells and their potential for immunotherapy. Immunol. Lett., 100, 42–55.
54) Iwabuchi, K. & Van Kaer, L. eds.(2019) Role of CD1- and MR1-restricted T cells in Immunity and Disease. pp.427, Frontiers Media (Lausanne).
55) Satoh, M., Namba, K.-i., Kitaichi, N., Endo, N., Kitamei, H., Iwata, D., Ohno, S., Ishida, S., Onoé, K., Watarai, H., et al. (2016) Invariant natural killer T cells play dual roles in the development of experimental autoimmune uveoretinitis. Exp. Eye Res., 153, 79–89.
56) Tajiri, N., Kato, T., Satoh, M., Iizuka, M., Taniguchi, M., Kitaichi, N., & Iwabuchi, K. (2021) The protective function of invariant natural killer T cells in the relapse of experimental autoimmune uveoretinitis. Exp. Eye Res., 203, 108406.
57) Sobirin, M.A., Kinugawa, S., Takahashi, M., Fukushima, A., Homma, T., Ono, T., Hirabayashi, K., Suga, T., Azalia, P., Takada, S., et al. (2012) Activation of natural killer T cells ameliorates postinfarct cardiac remodeling and failure in mice. Circ. Res., 111, 1037–1047.
58) Homma, T., Kinugawa, S., Takahashi, M., Sobirin, M.A., Saito, A., Fukushima, A., Suga, T., Takada, S., Kadoguchi, T., Masaki, Y., et al. (2013) Activation of invariant natural killer T cells by α-galactosylceramide ameliorates myocardial ischemia/reperfusion injury in mice. J. Mol. Cell. Cardiol., 62, 179–188.
59) Takahashi, M., Kinugawa, S., Takada, S., Kakutani, N., Furihata, T., Sobirin, M.A., Fukushima, A., Obata, Y., Saito, A., Ishimori, N., et al. (2020) The disruption of invariant natural killer T cells exacerbates cardiac hypertrophy and failure caused by pressure overload in mice. Exp. Physiol., 105, 489–501.
60) Saito, A., Ishimori, N., Tokuhara, S., Homma, T., Nishikawa, M., Iwabuchi, K., & Tsutsui, H. (2021) Activation of invariant natural killer T cells by α-galactosylceramide attenuates the development of angiotensin II-mediated abdominal aortic aneurysm in obese ob/ob mice. Front. Cardiovasc. Med., 8, 659418.
61) Takagi, D., Iwabuchi, K., Maeda, M., Nakamaru, Y., Furuta, Y., Fukuda, S., Van Kaer, L., Nishihira, J., & Onoé, K. (2006) Natural killer T cells ameliorate antibody-induced arthritis in macrophage migration inhibitory factor transgenic mice. Int. J. Mol. Med., 18, 829–836.
62) Ohmura, K., Ishimori, N., Ohmura, Y., Tokuhara, S., Nozawa, A., Horii, S., Andoh, Y., Fujii, S., Iwabuchi, K., Onoé, K., et al. (2010) Natural killer T cells are involved in adipose tissues inflammation and glucose intolerance in diet-induced obese mice. Arterioscler. Thromb. Vasc. Biol., 30, 193–199.
63) Satoh, M., Andoh, Y., Clingan, C.S., Ogura, H., Fujii, S., Eshima, K., Nakayama, T., Taniguchi, M., Hirata, N., Ishimori, N., et al. (2012) Type II NKT cells stimulate diet-induced obesity by mediating adipose tissue inflammation, steatohepatitis and insulin resistance. PLoS One, 7, e30568.
64) Satoh, M., Hoshino, M., Fujita, K., Iizuka, M., Fujii, S., Clingan, C.S., Van Kaer, L., & Iwabuchi, K. (2016) Adipocyte-specific CD1d-deficiency mitigates diet-induced obesity and insulin resistance in mice. Sci. Rep., 6, 28473.
65) Satoh, M., Iizuka, M., Majima, M., Ohwa, C., Hattori, A., Van Kaer, L., & Iwabuchi, K. (2022) Adipose invariant NKT cells interact with CD1d-expressing macrophages to regulate obesity-related inflammation. Immunology, 165, 414–427.
66) Diao, H., Kon, S., Iwabuchi, K., Kimura, C., Morimoto, J., Ito, D., Segawa, T., Maeda, M., Hamuro, J., Nakayama, T., et al. (2004) Osteopontin as a mediator of NKT cell function in T cell-mediated liver diseases. Immunity, 21, 539–550.
67) Wei, Y., Zeng, B., Chen, J., Cui, G., Lu, C., Wu, W., Yang, J., Wei, H., Xue, R., Bai, L., et al. (2016) Enterogenous bacterial glycolipids are required for the generation of natural killer T cells mediated liver injury. Sci. Rep., 6, 36365.
68) Nakai, Y., Iwabuchi, K., Fujii, S., Ishimori, N., Dashtsoodol, N., Watano, K., Mishima, T., Iwabuchi, C., Tanaka, S., Bezbradica, J.S., et al. (2004) Natural killer T cells accelerate atherogenesis in mice. Blood, 104, 2051–2059.
69) Andoh, Y., Ogura, H., Satoh, M., Shimano, K., Okuno, H., Fujii, S., Ishimori, N., Eshima, K., Tamauchi, H., Otani, T., et al. (2003) Natural killer T cells are required for lipopolysaccharide-mediated enhancement of atherosclerosis in apolipoprotein E-deficient mice. Immunobiology, 218, 561–569.
70) Caspi, R.R., Roberge, F.G., Chan, C.C., Wiggert, B., Chader, G.J., Rozenszajn, L.A., Lando, Z., & Nussenblatt, R.B. (1988) A new model of autoimmune disease. Experimental autoimmune uveoretinitis induced in mice with two different retinal antigens. J. Immunol., 140, 1490–1495.
71) Thurau, S.R., Chan, C.C., Nussenblatt, R.B., & Caspi, R.R. (1997) Oral tolerance in a murine model of relapsing experimental autoimmune uveoretinitis (EAU): induction of protective tolerance in primed animals. Clin. Exp. Immunol., 109, 370–376.
72) Kohno, H., Sakai, T., Tsuneoka, H., Imanishi, K., & Saito, S. (2009) Staphylococcal enterotoxin B is involved in aggravation and recurrence of murine experimental autoimmune uveoretinitis via Vβ8+CD4+ T cells. Exp. Eye Res., 89, 486–493.
73) Nieuwenhuis, E.E., Matsumoto, T., Exley, M., Schleipman, R.A., Glickman, J., Bailey, D.T., Corazza, N., Colgan, S.P., Onderdonk, A.B., & Blumberg, R.S. (2002) CD1d-dependent macrophage-mediated clearance of Pseudomonas aeruginosa from lung. Nat. Med., 8, 588–593.
74) Kinjo, T., Nakamatsu, M., Nakasone, C., Yamamoto, N., Kinjo, Y., Miyagi, K., Uezu, K., Nakamura, K., Higa, F., Tateyama, M., et al. (2006) NKT cells play a limited role in the neutrophilic inflammatory responses and host defense to pulmonary infection with Pseudomonas aeruginosa. Microbes Infect., 8, 2679–2685.
75) Hotamisligil, G.S., Shargill, N.S., & Spiegelman, B.M. (1993) Adipose expression of tumor necrosis factor-α: Direct role in obesity-linked insulin resistance. Science, 259, 87–91.
76) Xu, H., Barnes, G.T., Yang, Q., Tan, G., Yang, D., Chou, C.J., Sole, J., Nichols, A., Ross, J.S., Tartaglia, L.A., et al. (2003) Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest., 112, 1821–1830.
77) Nishimura, S., Manabe, I., Nagasaki, M., Eto, K., Yamashita, H., Ohsugi, M., Otsu, M., Hara, K., Ueki, K., Sugiura, S., et al. (2009) CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med., 15, 914–920.
78) Winer, S., Chan, Y., Paltser, G., Truong, D., Tsui, H., Bahrami, J., Dorfman, R., Wang, Y., Zielenski, J., Mastronardi, F., et al. (2009) Normalization of obesity-associated insulin resistance through immunotherapy. Nat. Med., 15, 921–929.
79) Feuerer, M., Herrero, L., Cipolletta, D., Naaz, A., Wong, J., Nayer, A., Lee, J., Goldfine, A.B., Benoist, C., Shoelson, S., et al. (2009) Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med., 15, 930–939.
80) Lumeng, C.N., Bodzin, J.L., & Saltiel, A.R. (2007) Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Invest., 117, 175–184.
81) Wu, D., Molofsky, A.B., Liang, H.E., Ricardo-Gonzalez, R.R., Jouihan, H.A., Bando, J.K., Chawla, A., & Locksley, R.M. (2011) Eosinophils sustain adipose alternatively activated macrophages associated with glucose homeostasis. Science, 332, 243–247.
82) Mantell, B.S., Stefanovic-Racic, M., Yang, X., Dedousis, N., Sipula, I.J., & O’Doherty, R.M. (2011) Mice lacking NKT cells but with a complete complement of CD8+ T-cells are not protected against the metabolic abnormalities of diet-induced obesity. PLoS One, 6, e19831.
83) Kotas, M.E., Lee, H.Y., Gillum, M.P., Annicelli, C., Guigni, B.A., Shulman, G.I., & Medzhitov, R. (2011) Impact of CD1d deficiency on metabolism. PLoS One, 6, e25478.
84) Wu, L., Parekh, V.V., Gabriel, C.L., Bracy, D.P., Marks-Shulman, P.A., Tamboli, R.A., Kim, S., Mendez-Fernandez, Y.V., Besra, G.S., Lomenick, J.P., et al. (2012) Activation of invariant natural killer T cells by lipid excess promotes tissue inflammation, insulin resistance, and hepatic steatosis in obsess mice. Proc. Natl. Acad. Sci. USA, 109, E1143–E1152.
85) Subramanian, S., Turner, M.S., Ding, Y., Goodspeed, L., Wang, S., Buckner, J.H., O’Brien, K., Getz, G.S., Reardon, C.A., & Chait, A. (2013) Increased levels of invariant natural killer T lymphocytes worsen metabolic abnormalities and atherosclerosis in obese mice. J. Lipid Res., 54, 2831–2841.
86) Ji, Y., Sun, S., Xu, A., Bhargava, P., Yang, L., Lam, K.S.L., Gao, B., Lee, C.-H., Kersten, S., & Qi, L. (2012) Activation of natural killer T cells promotes M2 macrophage polarization in adipose tissue and improves systemic glucose tolerance via the IL-4/STAT6 signaling axis in obesity. J. Biol. Chem., 287, 13561–13571.
87) Lynch, L., Nowak, M., Varghese, B., Clark, J., Hogan, A.E., Toxavidis, V., Balk, S.P., O’Shea, D., O’Farrelly, C., & Exley, M.A. (2012) Adipose tissue invariant NKT cells protect against diet-induced obesity and metabolic disorder through regulatory cytokine production. Immunity, 37, 574–587.
88) Schipper, H.S., Rakhshandehroo, M., van de Graaf, S.F., Venken, K., Koppen, A., Stienstra, R., Prop, S., Meerding, J., Hamers, N., Besra, G., et al. (2012) Natural killer T cells in adipose tissue prevent insulin resistance. J. Clin. Invest., 122, 3343–3354.
89) Strodthoff, D., Lundberg, A.M., Agardh, H.E., Ketelhuth, D.F., Paulsson-Berne, G., Arner, P., Hansson, G.K., & Gerdes, N. (2013) Lack of invariant natural killer T cells affects lipid metabolism in adipose tissue of diet-induced obese mice. Arterioscler. Thromb. Vasc. Biol., 33, 1189–1196.
90) Bedel, R., Matsuda, J.L., Brigl, M., White, J., Kappler, J., Marrack, P., & Gapin, L. (2012) Lower TCR repertoire diversity in Traj18-deficient mice. Nat. Immunol., 13, 705–706.
91) Ren, Y., Sekine-Kondo, E., Shibata, R., Kato-Itoh, M., Umino, A., Yanagida, A., Satoh, M., Inoue, K., Yamaguchi, T., Mochida, K., et al. (2017) A novel mouse model of iNKT cell-deficiency generated by CRISPR/Cas9 reveals a pathogenic role of iNKT cells in metabolic disease. Sci. Rep., 7, 12765.
92) Lynch, L., Michelet, X., Zhang, S., Brennan, P.J., Moseman, A., Lester, C., Besra, G., Vomhof-Dekrey, E.E., Tighe, M., Koay, H.F., et al. (2015) Regulatory iNKT cells lack expression of the transcription factor PLZF and control the homeostasis of T(reg) cells and macrophages in adipose tissue. Nat. Immunol., 16, 85–95.
93) Lynch, L., Hogan, A.E., Duquette, D., Lester, C., Banks, A., LeClair, K., Cohen, D.E., Ghosh, A., Lu, B., Corrigan, M., et al. (2016) iNKT cells induce FGF21 for thermogenesis and are required for maximal weight loss in GLP1 therapy. Cell Metab., 24, 510–519.
94) LaMarche, N.M., Kane, H., Kohlgruber, A.C., Dong, H., Lynch, L., & Brenner, M.B. (2020) Distinct iNKT cell populations use IFNγ or ER stress-induced IL-10 to control adipose tissue homeostasis. Cell Metab., 32, 243–258.
95) 日本食品標準成分表2020年版(八訂).文部科学省
96) Nieuwenhuis, E.E., Matsumoto, T., Lindenbergh, D., Willemsen, R., Kaser, A., Simons-Oosterhuis, Y., Brugman, S., Yamaguchi, K., Ishikawa, H., Aiba, Y., et al. (2009) Cd1d-dependent regulation of bacterial colonization in the intestine of mice. J. Clin. Invest., 119, 1241–1250.
97) Bai, L., Constantinides, M.G., Thomas, S.Y., Reboulet, R., Meng, F., Koentgen, F., Teyton, L., Savage, P.B., & Bendelac, A. (2012) Distinct APCs explain the cytokine bias of α-galactosylceramide variants in vivo. J. Immunol., 188, 3053–3061.
98) Wang, Z.V., Deng, Y., Wang, Q.A., Sun, K., & Scherer, P.E. (2010) Identification and characterization of a promoter cassette conferring adipocyte-specific gene expression. Endocrinology, 151, 2933–2939.
99) Clausen, B.E., Burkhardt, C., Reith, W., Renkawitz, R., & Förster, I. (1999) Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res., 8, 265–277.
100) Stranges, P.B., Watson, J., Cooper, C.J., Choisy-Rossi, C.M., Stonebraker, A.C., Beighton, R.A., Hartig, H., Sundberg, J.P., Servick, S., Kaufmann, G., et al. (2007) Elimination of antigen- presenting cells and autoreactive T cells by Fas contributes to prevention of autoimmunity. Immunity, 26, 629–641.
101) Zhang, H., Xue, R., Zhu, S., Fu, S., Chen, Z., Zhou, R., Tian, Z., & Bai, L. (2018) M2-specific reduction of CD1d switches NKT cell-mediated immune responses and triggers metaflammation in adipose tissue. Cell. Mol. Immunol., 15, 506–517.
102) Saxton, S.N., Heagerty, A.M., & Withers, S.B. (2020) Perivascular adipose tissue: An immune cell metropolis. Exp. Physiol., 105, 1440–1443.
103) Huh, J.Y., Park, J., Kim, J.I., Park, Y.J., Lee, Y.K., & Kim, J.B. (2017) Deletion of CD1d in adipocytes aggravates adipose tissue inflammation and insulin resistance in obesity. Diabetes, 66, 835–847.
104) Cui, S., Wang, C., Bai, W., Li, J., Pan, Y., Huang, X., Yang, H., Feng, Z., Xiang, Q., Fei, L., et al. (2020) CD1d1 intrinsic signaling in macrophages controls NLRP3 inflammasome expression during inflammation. Sci. Adv., 6, eaaz7290.
105) Toyoda, T., Kamata, T., Tanaka, K., Ihara, F., Takami, M., Suzuki, H., Nakajima, T., Ikeuchi, T., Kawasaki, Y., Hanaoka, H., et al. (2020) Phase II study of α-galactosylceramide-pulsed antigen-presenting cells in patients with advanced or recurrent non-small cell lung cancer. J. Immunother. Cancer, 8, e000316.
著者紹介Author Profile
 岩渕 和也(いわぶち かずや)
岩渕 和也(いわぶち かずや)北里大学名誉教授.医学博士.
略歴1956年札幌市に生る.82年北海道大学医学部卒業.86年同大学院博士課程修了,免疫科学研究所病理部門助手.89~91年米国ワシントン大学ハワードヒューズ医学研究所(HHMI@St.Louis)留学.99年北海道大学助教授.2010年北里大学医学部免疫学教授.18~20年同大学院医療系研究科長.22年同退職.
研究テーマと抱負生化学会誌にNKT細胞について書けることは望外の幸せですが,書きながらもう少し詳細なメカニズムを明らかに出来ていればと後悔頻りです.それらの解明は免疫学教室在籍の佐藤雅講師にバトンをパスしました.
ウェブサイトhttps://www.med.kitasato-u.ac.jp/lab/immunology/(在職時)
趣味料理.