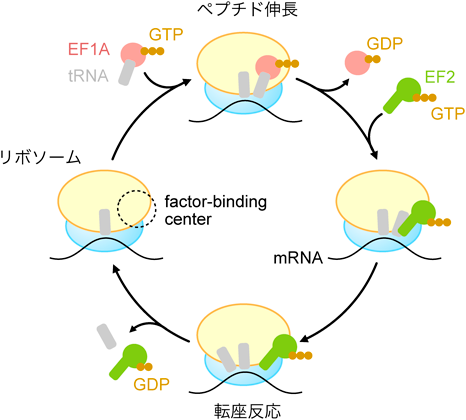
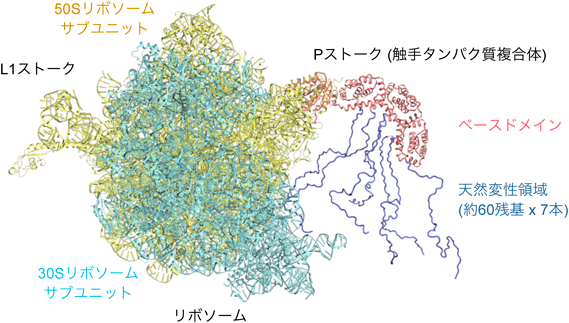
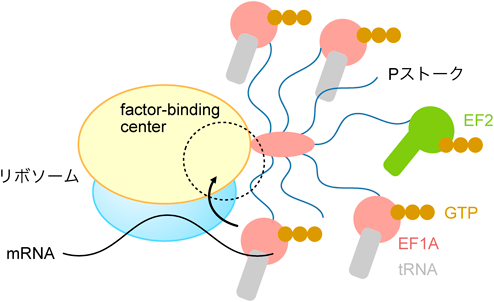
リボソームの触手による翻訳因子の収集機構Recruitment of translation factors by the tentacles of ribosomes
琉球大学大学院・医学研究科Graduate school of medicine, University of the Ryukyus ◇ 〒903–0215 沖縄県中頭郡西原町字上原207 ◇ 207 Uehara, Okinawa 903–0215, Japan
発行日:2022年10月25日Published: October 25, 2022