スプライシング阻害とRNA輸送Splicing inhibition and RNA transport
摂南大学農学部応用生物科学科Faculty of Agriculture, Setsunan University ◇ 〒573–0101 大阪府枚方市長尾峠町45–1 ◇ 45–1 Nagaotoge-cho, Hirakata, Osaka 573–0101, Japan
発行日:2022年12月25日Published: December 25, 2022
DNAから転写されたmRNA前駆体は,核内でキャップ,ポリAの付加,スプライシングを受ける.そして,成熟mRNAとなり,細胞質へと輸送され,翻訳の鋳型となる.mRNA前駆体はスプライシングが完了するまで核に繋留され細胞質へと輸送されることはない.この現象は酵母においてもヒトにおいても保存されている.しかし,その機構については,特に遺伝子の構造が複雑なヒトやマウスといった高等真核生物では不明な点が多い.
本稿では,古典的な酵母の遺伝学的的研究から,化学遺伝学的手法と次世代シーケンサーを組み合わせたアプローチを用いて得られた知見をもとにmRNA前駆体の核内繋留機構について概説する.
© 2022 公益社団法人日本生化学会© 2022 The Japanese Biochemical Society
転写,スプライシング,核外輸送,そして翻訳は独立したプロセスではなく,相互に密接に関連しておりその一部は共通の分子機構を有していることが知られている.一方で,スプライシング反応は,生命に必須の反応であり,その反応機構は複雑かつ冗長であるゆえに細胞内で反応を阻害することが困難だと考えられてきた.スプライシングを強力に阻害する低分子化合物が見いだされて以降,その特異的かつ強力な阻害作用により,スプライシングと,転写や核外輸送,そして翻訳との関連を証明する報告が相次いでいる.本稿では,そのなかでも主にスプライシングと核外輸送の関連に関する報告に絞り,筆者らが得た知見を交えて紹介する.
高等真核生物のほとんどの遺伝子にはイントロンがある.ヒトではmRNA前駆体1分子あたりイントロンが平均8.5個存在する.また,ヒトのエクソンの平均長は160塩基程度であるのに対し,イントロンの平均長は6100塩基にのぼる.出芽酵母では,mRNA前駆体1分子あたりイントロンは平均1.1個存在し,イントロンの平均長は310塩基であるので,出芽酵母に比べ高等真核生物の遺伝子がいかに複雑であるかがわかる.
イントロンはスプライシングによって正確に取り除かれる必要がある.イントロン内には5′スプライス部位(5′ss),3′スプライス部位(3′ss),そしてブランチ部位(BPS)と呼ばれるコンセンサス部位が存在する.これらコンセンサス部位の塩基配列は出芽酵母においては厳格に規定されているが,高等動物になるに従いコンセンサス配列の保存性は下がる.
スプライシングはスプライソソームと呼ばれる巨大なRNA–タンパク質複合体によって行われる.スプライソソームには5種類のU snRNP(small nuclear ribonucleoprotein particle),すなわちU1, U2, U5, U4/U6 snRNPと170種類以上ものタンパク質が存在する.U snRNPは100から190塩基程度のウリジンに富んだ低分子RNA(U snRNA)であり,U snRNPのRNA成分とmRNA前駆体との間のRNAどうしの塩基対合がスプライス部位の認識に必要である.
スプライソソームの形成は段階的に行われる.第2段階の反応を経た成熟mRNAは,核膜孔複合体を経て細胞質へと輸送され翻訳の鋳型となる.一方でスプライシングが未完了のmRNA前駆体はスプライシングが完了するまで核に繋留される(詳細は片岡の稿参照).
mRNA前駆体が細胞質へ漏出し,細胞質でイントロンが翻訳されると,イントロン内に含まれるナンセンスコドンにより天然には存在しない短縮型異常タンパク質が産生される恐れがある.通常ナンセンス変異を持つ異常なmRNAは,正常なmRNAと異なる経路,ナンセンス変異依存分解(NMD)を経て急速に分解される.したがって細胞質に漏出したmRNA前駆体もナンセンス変異を持つため異常なmRNAとして認識されるはずである.しかし,後述するが,スプライシング阻害を受けた細胞において,NMDによる分解を免れるmRNA前駆体も多数確認されている1).
また,イントロンにはSINEやLINEなどといったリピート関連配列が豊富に存在するため,しばしばリピート配列どうしの塩基対結合により二次構造をとることがあり,それらが細胞質に存在するとPKR・RLR・TLRといった二本鎖RNA(dsRNA)を感知する自然免疫系のセンサーを活性化させ,翻訳抑制等のウイルス応答を引き起こす(岩崎らの稿を参照)2).したがって不要なウイルス応答を抑えるためにもmRNA前駆体は核内の転写部位に繋留され,迅速にスプライシングが行われる必要がある.
同様に,スプライシング反応の完了後,切り出されたラリアット型イントロンRNAもリピート配列どうしの塩基対結合によりdsRNAを形成しうる.ラリアット型イントロンは,環状構造を持つゆえに,mRNA前駆体よりさらに強固な二次構造をとることが予想される3).したがってラリアット型イントロンの細胞質への漏出も防ぐ必要があり,一部の例外を除いて,ラリアット型イントロンRNAも核内に繋留され,迅速に分解される4).
出芽酵母を用いた分子遺伝学的研究により,イントロン配列を持つmRNA前駆体を核に繋留するために必要な因子の探索が行われてきた.たとえば,スプライシング因子であるBBP/SF1やSF3Bの温度感受性変異株において,スプライシングの阻害によって過剰に安定化されたmRNA前駆体が細胞質へと漏出することが示されている5, 6).また,核膜孔複合体の構成成分であるMlp1の欠損株は,mRNA前駆体の安定化を伴うことなくmRNA前駆体の細胞質への漏出を引き起こし,さらにスプライシング因子Prp18とMlp1の二重欠損株は合成致死となることから,Mlp1は核膜孔複合体付近でmRNA前駆体の漏出を監視していることが示唆されている7).
高等真核生物においても,mRNA前駆体の核内繋留機構が報告されている.32P標識したin vitroで合成したmRNA前駆体とアフリカツメガエル卵母細胞とを用いる核外輸送アッセイはRNAの輸送を研究する上で現在でも非常に強力なツールである.エクソン配列にしばしば出現するESE(exonic splicing enhancer)配列は,スプライシングを促進するという本来の活性以外に,mRNA前駆体を核にとどめておく活性がある8).興味深いことにESEによる核内繋留効果はスプライシングが完了すると消失する.このことはスプライシング反応の完了がmRNA前駆体の核内繋留機構の解除の引き金になっていることを示唆する.またsiRNAでスプライシング因子をノックダウンした培養細胞とレポーター遺伝子を用いた実験から,スプライシング因子であるU1 snRNPとU2AF65がmRNA前駆体の核内繋留因子として同定されている9).
しかし前述のとおり,高等真核生物では,酵母に比べて遺伝子のイントロンは長大で,複数存在する.したがって,mRNA前駆体の核内繋留機構を調べる上で,合成RNAやレポーター遺伝子を用いた実験には限界が生じる.また,siRNAを用いた培養細胞のノックダウン実験では24~72時間かけて標的遺伝子の発現を徐々に抑制するため,RNAの局在に関して明確な結果が出にくいなどの課題がある.そこで,mRNA前駆体のスプライシングや核内繋留機構を強力に阻害する低分子化合物が待ち望まれていた.
天然物由来の低分子化合物であるFR901464はもともと強力な制がん作用を持つ化合物として同定されたが,その作用機序は不明であった10–12).甲斐田らは,FR901464にメチル基を導入したより安定な化合物であるスプライソスタチンA(SSA)がU2 snRNPの構成因子であるSF3B1に直接結合することによってスプライシング反応を強力に阻害することを見いだした13).以降,天然物および合成誘導体のSF3B1をターゲットとするスプライシング阻害化合物が次々と見いだされているのでここに列挙しておく(図1)14).ここでは詳細は述べないが,X線結晶解析やクライオ電子顕微鏡解析からスプライソスタチン類・プラジエノライド類はいずれもU2 snRNPの構成因子であるSF3B1とPHF5Aが作るブランチ部位認識ポケットに入り込み,mRNA前駆体に対して競合的に阻害することが示されている.一方で,スプライソスタチン類のうち,C3位にエポキシ基を持つものは,U2 snRNPの構成因子であるPHF5Aの26番目のシステイン残基と共有結合をとる.この共有結合はプラジエノライド類には起こらないことなどからスプライソスタチン類・プラジエノライド類の阻害様式は完全に同一ではないようである.
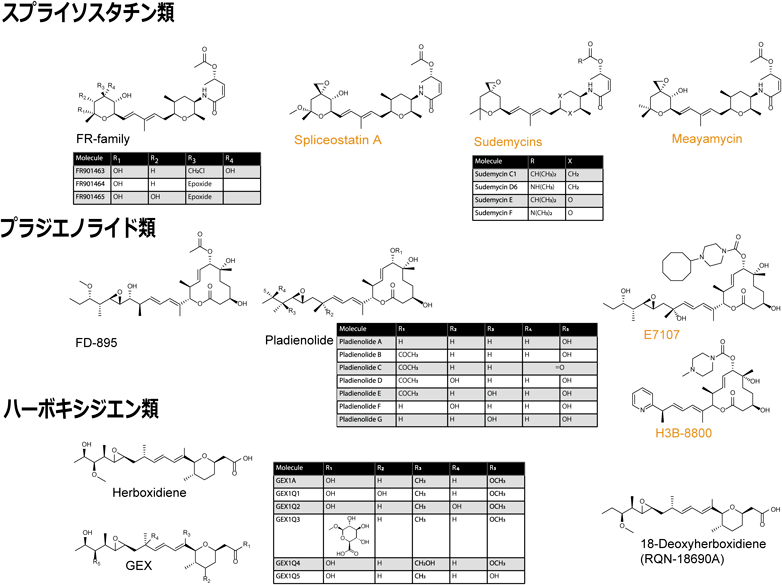
元の天然物の合成および半合成誘導体をオレンジ色で示した.J. Antibiot., 74, 603–616 (2021)14) よりCC BY 4.0のもと転載.
甲斐田らは,p27遺伝子の上流のイントロンにHAタグを,下流のエクソンにMycタグを導入したミニジーンを作製した(p27-intHA).スプライシングされたmRNAが翻訳されるとMycタグで標識されたタンパク質が産生されるが,mRNA前駆体が翻訳されるとHAタグで標識されたタンパク質が産生される.甲斐田らはこのレポーターを用いて,SSAによるスプライシング阻害がp27 mRNA前駆体のスプライシングを阻害するばかりではなく,p27 mRNA前駆体の翻訳を引き起こすことを明らかにした13).
著者らは,ゲノムワイドにmRNA前駆体の核内繋留機構を調べる目的で,SSA処理したヒトHeLa S3細胞の核・細胞質を分画し,RNA-Seqを行った.RNA-SeqはcDNAを短く分解して高速シーケンサーで配列を決定する方法であるが,遺伝子発現量を推定するばかりでなく,選択的スプライシングを解析することも可能である.一般に,選択的スプライシングには五つのパターンがあるが,SSAは主にイントロン保持によりmRNA前駆体の安定化を引き起こす(岩崎らの稿参照).SSA処理によりイントロン保持されやすいイントロン配列を解析したところ,通常のイントロンに比べて弱いブランチ部位(BPS)を持っていた.このことはSSAがSF3Bに結合することで,SF3Bのブランチ部位の認識を弱めることによりスプライシング反応を阻害するという事実と一致する15).
また,細胞質画分にて有意に出現するmRNA前駆体の特徴を調べたところ,弱いBPSを持つことに加えて(i)弱い5′ ssを持つ,(ii)遺伝子長が短い,(iii)エクソン(イントロン)数が少ない,という特徴が浮かび上がってきた.一方で遺伝子長が長いmRNA前駆体は細胞質へ漏出することなく,核内のクロマチン画分に繋留された.
以上の結果より核内繋留機構のモデルとして以下のモデルが推測される(図2).塩基長が短いmRNA前駆体は,転写に要する時間が短時間であるため,転写が終了した後にスプライシングが行われる(post-transcriptional splicing).したがってmRNA前駆体がスプライソソームに取り込まれることによって核内に繋留され,スプライシングの完了を待って核外輸送される.
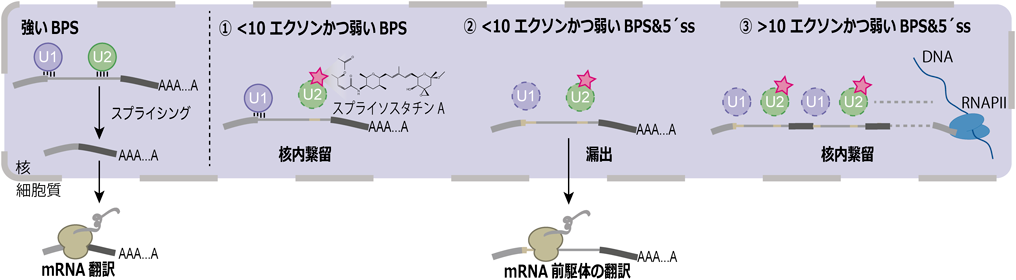
一方で,長大な,すなわちエクソン数が多い遺伝子のスプライシングは転写に長時間を要するため,それら長大な遺伝子内のほとんどのイントロンは転写と共役する形でスプライシングを受ける(co-transcriptional splicing).高等真核生物では,長大な遺伝子のスプライシングを正確に行う必要があるため,転写と共役する形でスプライシングが起こる方向に進化していったのであろう.RNAポリメラーゼIIを中心とした転写装置につながったmRNA前駆体はスプライシングが完了するまで転写部位にとどめられると考えられる.mRNA前駆体の翻訳物が細胞に与える影響については,質量分析法やリボソームプロファイリング法を用いて詳細に解析されている(岩崎らの稿を参照).
DNAから転写されるRNAには,mRNAの他に,タンパク質をコードしない非コードRNA(ncRNA)がある.ncRNAにはマイクロRNA(miRNA),リボソームRNA(rRNA),U snRNAなどの他,200塩基以上の長鎖非コードRNA(lncRNA)がある.lncRNAは転写・スプライシング・エピジェネティクスの制御を介して,発生や増殖に関わっている一方で,ncRNAの異常はがんや精神疾患などをもたらすことがわかってきている.lncRNAの多くは核に局在し,スプライシングを受けない.その核局在メカニズムは不明であったが近年スプライシング因子との関連が示唆されている16, 17).そこで著者らは,SSA処理細胞のRNA-Seqの再解析を行い,lncRNAの局在の解析を行った.その一つであるMALAT1は核内での著しい減少と細胞質での増加がみられた.MALAT1は予後の悪い非小細胞肺がんにおいて発現上昇がみられるlncRNAとして見いだされ,主に核スペックルに局在する18).
通常のmRNA前駆体はイントロンが取り除かれて成熟すると,不要な末端が切断され,そこにアデノシン(A)が複数連続したポリA配列が付加される.しかし,本来のポリA付加部位より上流のポリA付加部位にて切断とポリA付加が起きることがある.これを「未成熟切断およびポリA付加(PCPA)」という(岩崎らの稿参照).通常PCPAは,U1 snRNPによって阻害されている(図3C).MALAT1は,スプライシングもポリA付加も受けない長いRNA(約8 kb)として存在するが,SSAを添加するとPCPAが引き起こされ,短いMALAT1のアイソフォーム(約6 kbのポリA付加を受けたRNA)を生じる(図3D).
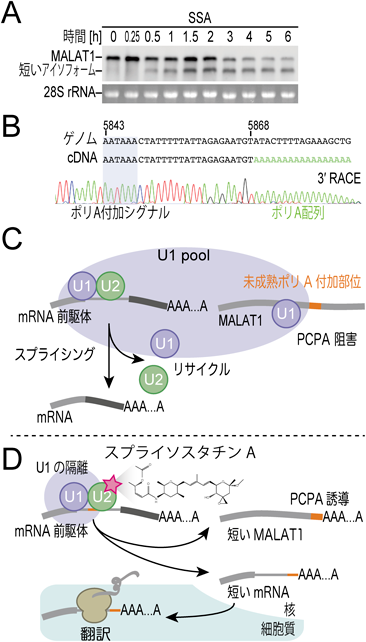
(A)スプライソスタチンA(SSA)処理細胞におけるMALAT1 RNAの経時変化.SSAの処理時間に応じて短いアイソフォームが出現する.(B) MALAT1の短いアイソフォームの末端配列を調べた.下流の末端にはゲノムには存在しないポリA配列(緑)が含まれることから,上流にあるポリA付加シグナル(AAUAAA)が認識され,未成熟切断およびポリA付加(PCPA)が起こっていることがわかった.(C)通常,mRNA前駆体はイントロンが取り除かれて成熟すると,不要な末端が切断され,そこにポリAが付加される.未成熟切断およびポリA付加(PCPA)は,スプライング因子U1によって阻害されている.MALAT1は,スプライシングもポリA付加も受けない長鎖非コードRNAとして核内に存在する.(D)スプライソスタチンA(SSA)を添加すると,スプライシング因子のU2に結合し(赤星印),U2が機能不全となる.すると機能不全U2がmRNA前駆体–U1–U2複合体を安定化し,U1とU2のリサイクルが阻害される.これにより,核内の利用可能なU1の濃度が不足し,PCPAを抑えきれなくなってMALAT1は切断され,ポリA付加される.これとは別に,核内では短いRNAが生じ,それが細胞質へ移行して,新奇タンパク質が合成されることもわかった.
次に,著者らはSSAがPCPAを引き起こすメカニズムについて詳細に調べた.その結果,(1)SSAはU2内のSF3bに結合することで機能不全U2を生じること,(2)機能不全U2がmRNA前駆体–U1–U2複合体を安定化することでスプライシング因子のリサイクルを阻害すること,(3)リサイクル阻害により核内の利用可能なU1の濃度が不足し,PCPAを抑えきれなくなってMALAT1は切断され,ポリA付加が引き起こされることが明らかになった(図3D).U1 snRNPはMALAT1のPCPAを抑制する他に,MALAT1の核内への局在にも関与すると報告されている16).また,多くの動物種においてMALAT1のポリA部位が保存されている.MALAT1のポリA部位には,まだ知られていない重要な機能があるのかもしれない.
最後に,MALAT1以外にSSAによってPCPAが誘発されるRNA種を網羅的に探索した.その結果,mRNAの一部において,主に第1イントロン中の潜在的ポリA付加部位でのPCPAが起こり,第1エクソンとイントロンからなる短いRNAが生じて細胞質へと輸送されていることが明らかになった(図3D).さらに,それらがタンパク質に翻訳されるかどうかをリボソームプロファイリング法によって網羅的に解析したところ,この短いRNAの翻訳がみられたことから,SSA処理した細胞の中では,イントロン由来の配列を持った新奇タンパク質が生じていることがわかった(図3D).
本稿で紹介したとおり,著者らは天然物由来のスプライシング調節化合物が持つ強力な生理活性に導かれてmRNA前駆体の核内繋留機構やスプライシング因子のリサイクルの意義を理解することができた.近年1細胞核・細胞質分画RNAシーケンス法が確立されている19).こういった技術を組み合わせてさまざまな組織の細胞に含まれるmRNA前駆体の挙動を調べることで未知の発見がもたらされるに違いない.
1) Yoshimoto, R., Kaida, D., Furuno, M., Burroughs, A.M., Noma, S., Suzuki, H., Kawamura, Y., Hayashizaki, Y., Mayeda, A., & Yoshida, M. (2017) Global analysis of pre-mRNA subcellular localization following splicing inhibition by spliceostatin A. RNA, 23, 47–57.
2) Bowling, E.A., Wang, J.H., Gong, F., Wu, W., Neill, N.J., Kim, I.S., Tyagi, S., Orellana, M., Kurley, S.J., Dominguez-Vidana, R., et al. (2021) Spliceosome-targeted therapies trigger an antiviral immune response in triple-negative breast cancer. Cell, 184, 384–403.e21.
3) Liu, C.X., Li, X., Nan, F., Jiang, S., Gao, X., Guo, S.K., Xue, W., Cui, Y., Dong, K., Ding, H., et al. (2019) Structure and Degradation of Circular RNAs Regulate PKR Activation in Innate Immunity. Cell, 177, 865–880.e21.
4) Yoshimoto, R., Kataoka, N., Okawa, K., & Ohno, M. (2009) Isolation and characterization of post-splicing lariat-intron complexes. Nucleic Acids Res., 37, 891–902.
5) Rutz, B. & Seraphin, B. (2000) A dual role for BBP/ScSF1 in nuclear pre-mRNA retention and splicing. EMBO J., 19, 1873–1886.
6) Dziembowski, A., Ventura, A.P., Rutz, B., Caspary, F., Faux, C., Halgand, F., Laprevote, O., & Seraphin, B. (2004) Proteomic analysis identifies a new complex required for nuclear pre-mRNA retention and splicing. EMBO J., 23, 4847–4856.
7) Galy, V., Gadal, O., Fromont-Racine, M., Romano, A., Jacquier, A., & Nehrbass, U. (2004) Nuclear retention of unspliced mRNAs in yeast is mediated by perinuclear Mlp1. Cell, 116, 63–73.
8) Taniguchi, I., Masuyama, K., & Ohno, M. (2007) Role of purine-rich exonic splicing enhancers in nuclear retention of pre-mRNAs. Proc. Natl. Acad. Sci. USA, 104, 13684–13689.
9) Takemura, R., Takeiwa, T., Taniguchi, I., McCloskey, A., & Ohno, M. (2011) Multiple factors in the early splicing complex are involved in the nuclear retention of pre-mRNAs in mammalian cells. Genes Cells, 16, 1035–1049.
10) Nakajima, H., Takase, S., Terano, H., & Tanaka, H. (1997) New antitumor substances, FR901463, FR901464 and FR901465. III. Structures of FR901463, FR901464 and FR901465. J. Antibiot. (Tokyo), 50, 96–99.
11) Nakajima, H., Hori, Y., Terano, H., Okuhara, M., Manda, T., Matsumoto, S., & Shimomura, K. (1996) New antitumor substances, FR901463, FR901464 and FR901465. II. Activities against experimental tumors in mice and mechanism of action. J. Antibiot. (Tokyo), 49, 1204–1211.
12) Nakajima, H., Sato, B., Fujita, T., Takase, S., Terano, H., & Okuhara, M. (1996) New antitumor substances, FR901463, FR901464 and FR901465. I. Taxonomy, fermentation, isolation, physico-chemical properties and biological activities. J. Antibiot. (Tokyo), 49, 1196–1203.
13) Kaida, D., Motoyoshi, H., Tashiro, E., Nojima, T., Hagiwara, M., Ishigami, K., Watanabe, H., Kitahara, T., Yoshida, T., Nakajima, H., et al. (2007) Spliceostatin A targets SF3b and inhibits both splicing and nuclear retention of pre-mRNA. Nat. Chem. Biol., 3, 576–583.
14) Schneider-Poetsch, T., Chhipi-Shrestha, J.K., & Yoshida, M. (2021) Splicing modulators: on the way from nature to clinic. J. Antibiot. (Tokyo), 74, 603–616.
15) Corrionero, A., Minana, B., & Valcarcel, J. (2011) Reduced fidelity of branch point recognition and alternative splicing induced by the anti-tumor drug spliceostatin A. Genes Dev., 25, 445–459.
16) Yin, Y., Lu, J.Y., Zhang, X., Shao, W., Xu, Y., Li, P., Hong, Y., Cui, L., Shan, G., Tian, B., et al. (2020) U1 snRNP regulates chromatin retention of noncoding RNAs. Nature, 580, 147–150.
17) Lubelsky, Y. & Ulitsky, I. (2018) Sequences enriched in Alu repeats drive nuclear localization of long RNAs in human cells. Nature, 555, 107–111.
18) Yoshimoto, R., Mayeda, A., Yoshida, M., & Nakagawa, S. (2016) MALAT1 long non-coding RNA in cancer. Biochim. Biophys. Acta, 1859, 192–199.
19) Abdelmoez, M.N., Iida, K., Oguchi, Y., Nishikii, H., Yokokawa, R., Kotera, H., Uemura, S., Santiago, J.G., & Shintaku, H. (2018) SINC-seq: correlation of transient gene expressions between nucleus and cytoplasm reflects single-cell physiology. Genome Biol., 19, 66.

摂南大学農学部 講師.理学博士.
1979年長崎県に生まれる.2002年京都大学薬学部卒業.理化学研究所ポスドク・藤田医科大学助教を経て20年より現職.
研究テーマと抱負mRNAスプライシングに関わるノンコーディングRNAの機能解析.大学院生の頃はin vitroスプライシング反応系を駆使した生化学的な研究からはじめ,近年はbioinformaticsの力を借りている.「一生に一度くらいは大当たり」を目指して日々精進.
ウェブサイトhttps://www.researchgate.net/profile/Rei-Yoshimoto
趣味読書,DIY.

This page was created on 2022-11-16T16:36:35.67+09:00
This page was last modified on 2022-12-14T11:18:54.000+09:00
このサイトは(株)国際文献社によって運用されています。