スプライシング異常と細胞周期停止機構Cell cycle arrest caused by splicing abnormality
富山大学学術研究部医学系Faculty of Medicine, Academic Assembly, University of Toyama ◇ 〒930–0194 富山県富山市杉谷2630 ◇ Sugitani 2630, Toyama 930–0194, Japan
発行日:2022年12月25日Published: December 25, 2022
真核生物において,mRNAスプライシングは遺伝子発現にとって必須のメカニズムである.これまでの研究から,スプライシング関連因子の変異やスプライシング阻害剤は,細胞周期関連因子のスプライシングを阻害し,遺伝子発現を低下させることで細胞周期停止を引き起こすことが明らかとなった.さらには,スプライシング阻害により蓄積したpre-mRNAが翻訳されたトランケート型タンパク質が細胞周期停止に関わることもわかってきた.近年,スプライシング阻害剤は新規の作用機序を有する強力な抗がん剤として注目を浴びており,これらの細胞周期停止メカニズムの解明は,スプライシング阻害剤を基盤とした抗がん剤の開発を強力に後押しすると考えられる.本稿では,mRNAスプライシング異常と細胞周期停止に関するこれまでの知見を紹介する.
© 2022 公益社団法人日本生化学会© 2022 The Japanese Biochemical Society
真核生物において,mRNAスプライシングは遺伝子発現にとって必須のメカニズムである1–3).特にヒトにおいては,遺伝子全体の95%以上がスプライシングを受けるため,スプライシングの異常は,ほぼすべての遺伝子発現,ならびに細胞機能に影響を与えるといっていい.当然,スプライシング異常や低分子化合物によるスプライシング阻害は細胞周期進行にも影響を与える4).一方,スプライシング阻害剤は非常に強力な抗がん活性を有することが報告されており,骨髄異形成症候群や慢性リンパ性白血病などのいわゆる血液のがん,さらには種々の固形がんに対しても臨床研究が進められている5–11).しかしながら,抗がん活性を発揮する作用機序には未知の部分も多く,この解明がスプライシング阻害剤を基盤とした抗がん剤の開発を強力に後押しすると考えられる.本稿では,抗がん活性の原因の一つであると考えられる,mRNAスプライシングの異常と細胞周期停止に関して概説したい.
細胞周期進行は非常に厳密に制御されており,多くの因子が関わっている(図1).細胞周期を進行させる役割を果たす因子として,サイクリンやサイクリン依存性キナーゼ(以下CDK)がよく知られており,これらは細胞周期進行のエンジンとして働いている12).細胞周期進行に関わるサイクリンタンパク質としては,サイクリンA, B, D, Eなどが知られており,それぞれ細胞周期の異なるタイミングで発現が誘導され,機能を果たしている.これらのサイクリンがパートナーであるCDKと結合し,基質を適切なタイミングでリン酸化することで,正確に細胞周期が進行していく.たとえば,サイクリンDはCDK4/6と複合体を作ることで,G1期の進行に寄与する.サイクリンEはG1期の後半からS期の前半にかけて発現が確認され,CDK2と複合体を形成することにより,G1期からS期への移行に重要な役割を果たしている.サイクリンAはS期の初めから発現が確認され,M期へと移行する際に急激に減少していく.サイクリンAは,S期ではCDK2と複合体を形成し正確なS期の進行をコントロールしている.S期の終盤になると,サイクリンAはCDK1と複合体を形成し,G2期からM期への移行を制御する.サイクリンBは主にG2期からM期にかけて発現が確認され,CDK1と複合体を形成することによりM期の開始に重要な役割を果たす.一方,M期からG1期への移行には阻害的に働くため,M期の終盤になると分解されその役割を終える.
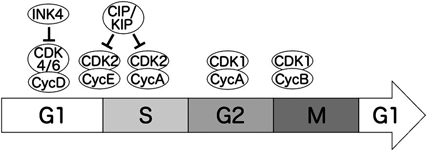
細胞周期により異なる因子が細胞周期進行を制御している.
一方,ブレーキとして働くのはCDKインヒビターである13).CDKインヒビターは,CDKタンパク質に結合し,その機能を阻害することにより細胞周期進行を抑制している.CDKインヒビターはINK4とCIP/KIPという二つのグループに分けられる.INK4に属するのはp15, p16, p18, p19という四つのタンパク質であり,これらはCDK4/6と結合しサイクリンDとの結合を阻害することで,CDK4/6の活性を抑制する.一方,CIP/KIPにはp21, p27, p57が含まれ,これらはCDK2-サイクリンA/E複合体と結合することで,G1期からS期にかけての制御を行っている.
siRNAを用いたゲノムワイドなスクリーニングにより,多くの細胞周期進行に関わる遺伝子が同定された.非常に興味深いことに多くのスプライシング因子も細胞周期関連遺伝子として同定された14).また,個別の研究からも,細胞周期進行に関わるスプライシング因子が多数同定されており,スプライシングと細胞周期進行には密接な関係があることが予想される15–19).ここでは,細胞周期進行に関わるいくつかのスプライシング関連因子とそのメカニズムを紹介する.
選択的スプライシングは,スプライシングの際にイントロンとして切り出す配列を変えることにより,一つの遺伝子から複数種類のアイソフォームを作り出す仕組みである.細胞周期により選択的スプライシングのパターンが変化する遺伝子を探索したところ,約1300遺伝子が細胞周期に従って異なるアイソフォームを作ることが明らかとなった20).また,これらの多くは細胞周期進行に関わる遺伝子であった.細胞周期依存的な選択的スプライシングの変化に関わる因子を探索したところ,選択的スプライシングに関わる因子の一つであるCLK1の発現が細胞周期の進行に従って変動することが明らかとなった.実際に,1300遺伝子のうち約半数は,CLK1により選択的スプライシングが制御されていた.このような結果から,CLK1が細胞周期進行に関与することが予想される.実際CLK1をノックダウンすることにより細胞周期がS期からG2期で停止し,細胞増殖が抑えられることが明らかとなった.一方,がん細胞では正常細胞と比較してCLK1の発現が高く,CLK1の発現レベルが高いほど予後も悪いという結果も出ている.これらのことから,CLK1は細胞周期進行を正に制御し,その異常な高発現は選択的スプライシングのパターンを変化させ,がんの悪性度に影響すると考えられる.
SRSF1は,選択的スプライシングの制御因子であり,がん細胞ではその過剰発現が観察される21).一方,SRSF1のノックダウンにより細胞周期がG2期で停止し,アポトーシスが引き起こされる22).このように,SRSF1は細胞周期進行やがん化と密接な関係があることが知られている.また,SRSF1は活性や局在などがリン酸化により制御されることも知られている23).SRSF1はC末端側にセリンとアルギニンに富んだ領域を持ち,この領域がリン酸化されることが活性化に重要であると考えられているが,N末端側にあるRNA結合部位中にある19番目のチロシン(Tyr-19)のリン酸化が,急性リンパ芽球性白血病のサンプルで亢進していることが明らかとなった24).Tyr-19のリン酸化は細胞内局在の変化を引き起こし,また,Tyr-19をアスパラギン酸に置換したリン酸化ミミック体の過剰発現は細胞増殖を促進した.一方,リン酸化を受けないフェニルアラニンに置換した変異体の過剰発現によりG1期の細胞が増加した.これらのことから,Tyr-19のリン酸化が細胞周期進行に重要な働きをしていると考えられる.今後の研究により,このリン酸化により引き起こされる細胞周期関連因子のスプライシングパターンの変化など,さらなるメカニズムの解明が期待される.
Prp19複合体は,スプライソソームに関わる因子として同定されたが,その後の研究から転写,ユビキチン化,DNAダメージレスポンスなどにも関わる多機能な因子であることも明らかとなった25).CDC5はPrp19複合体の構成因子であり,CDC5自身がスプライシング反応に必須であることが報告されている26, 27).また,CDC(cell division cycle)5は,その名のとおり細胞周期進行に必要な因子であり,CDC5変異体の過剰発現やCDC5のノックダウンにより細胞周期がG2/M期で停止する28–30).また,CDC5のノックダウンにより,M期進行に関わる遺伝子のスプライシングが阻害され,発現が低下することも明らかとなった28).これらのことから,CDC5はM期進行に関わる遺伝子のスプライシングを介して細胞周期を制御していると考えられる.興味深いことに,CDK2-サイクリンEによりリン酸化されたCDC5がスプライソソームの構成因子の一つであるNIPPと結合することから,細胞周期依存的なリン酸化が細胞周期関連遺伝子のスプライシングや遺伝子発現に関わるのかもしれない31–33).実際に,出芽酵母ではα-チューブリン遺伝子のイントロンを取り除くことにより,CDC5の変異による細胞周期停止が回復することもわかっている34).ただし,出芽酵母はイントロンを有する遺伝子がきわめて少ないため,1遺伝子のスプライシング異常の問題を解決するだけで,細胞周期を回復させることができるが,高等真核生物ではほとんどの遺伝子がスプライシングを受けるため,このようなアプローチでは細胞周期を回復させることはできないだろう.
上述のように,スプライシング因子の変異やsiRNAによるノックダウンで細胞周期に影響が出ることが明らかとなっている.しかしながら,高等真核生物の場合,スプライシングはほぼすべての遺伝子発現に関与するため,siRNAのようにノックダウンまでに時間がかかる手法では二次的,三次的な効果を観察している可能性もある.一方,近年発見されたスプライシング阻害剤を用いることにより,スプライシング阻害の直後の細胞のようすを観察することにより,より直接的な影響を観察できると考えられる.ここからは,そのようなスプライシング阻害剤を用いた研究を紹介したい.
スプライソスタチンA(以下SSA),プラジエノライドB(以下Pla-B),ハーボキシジエンなどのスプライシング阻害剤は,強い抗がん活性を持ち,またG1期およびG2/M期において細胞周期を停止させることが報告されてきた5–8, 35).また,SSA処理細胞中では,スプライシング阻害により蓄積したpre-mRNAが蓄積され,イントロン部分が翻訳されたp27 CDKインヒビターのC末端トランケート型タンパク質(以下p27*)が高発現していることも明らかとされたものの,p27*と細胞周期停止の関係は明らかにはなっていなかった5).p27はスプライシングを受けたCDKN1B mRNAから翻訳され産生されるが,スプライシング阻害時にはイントロン1が残ったCDKN1B pre-mRNAが蓄積し,それが翻訳されることでp27*が産生される(図2).p27全体は198アミノ酸であるが,そのうちエクソン1由来のアミノ酸配列が全体の約8割を占める.したがって,p27*はその大部分がp27と同一のアミノ酸配列であり,N末端側にあるCDK結合ドメインなども有している.実際に,p27, p27*のいずれもCDK2-サイクリンE複合体と結合し,そのリン酸化活性を阻害することが知られている.
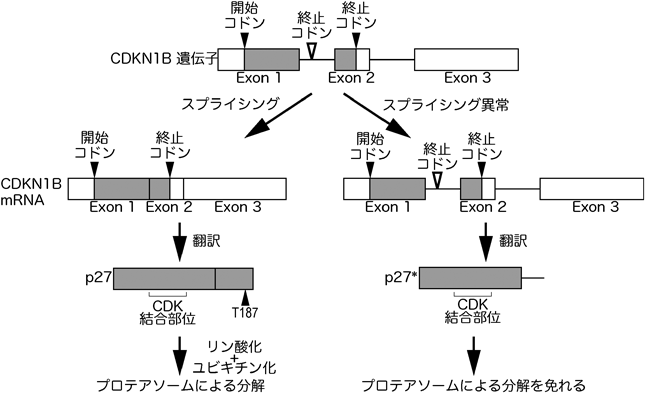
CDKN1B pre-mRNAがスプライシングを受けたのちに翻訳されるとp27が生成され,スプライシングが阻害されpre-mRNAが翻訳されるとp27*が生成される.p27には,C末端にリン酸化サイトがあり,このサイトがリン酸化されることにより,ユビキチン化され,プロテアソームにより分解される.一方,p27*にはこのリン酸化サイトが存在しない.
それでは,p27*が作られることが,スプライシング阻害によるG1期停止の原因なのだろうか? 培養細胞中でp27*を過剰発現させることにより,G1期で停止する細胞が増加することが報告されている36).また,p27*過剰発現によりG1期停止を示す細胞の割合は,p27過剰発現によるものと同程度であるため,CDK2-サイクリンE複合体に対する阻害効果や,G1期停止を引き起こす能力に関してはp27とp27*に差はないと考えられる.しかしながら,これらの因子を過剰発現しても完全なG1期停止は観察されないため,スプライシング阻害によるG1期停止にはp27*以外の因子も関与していると考えられる.それを裏づけるように,p27およびp27*ノックダウン細胞をスプライシング阻害剤で処理した場合でも,G1期停止する細胞の割合は低下するものの,一部の細胞はG1期で停止する.これらのことから,スプライシング阻害によるp27*の産生は,G1期停止の原因の一つであるものの,それだけでは説明できない部分も残されていると考えられる.
上述のように,p27*の発現だけではスプライシング阻害剤によるG1期停止は説明できない.それでは,G1期からS期への移行に関わる他の因子には変化がみられるのだろうか? きわめて多くの因子がこの過程に関わっていることが知られているが,その中でもサイクリンEやE2F1のタンパク質レベルがスプライシング阻害により顕著に減少することが明らかとなった37).一方,サイクリンEのパートナーであるCDK2のタンパク質量には明確な変化は観察されなかったことから,特定の因子の減少がG1期停止を引き起こしていると考えられる.また,これらのタンパク質の減少は,mRNAレベルの減少で説明できることも報告された.それでは,これらのmRNAはどのようなメカニズムで減少するのであろうか? 一般的に,スプライシングを受けていないpre-mRNAが蓄積すると,異常なmRNAであると見なされ,核内のエキソソームによる分解や,細胞質でのnonsense mediated mRNA decayなどのメカニズムにより分解されることが知られている.しかしながら,これらのmRNAはこれらの分解機構による分解を受けていないことが明らかとなった.さらに,これらの遺伝子の転写開始には問題がないものの,転写伸長ができず,結果的にmRNAの全長が転写されないことがmRNAレベルの低下を引き起こしていることも明らかとなった.スプライソソームの構成因子であるU1 snRNPの阻害により,転写途中のpre-mRNAが本来のポリA位置より上流で異常なポリA化を受け,転写が早期に集結してしまうことや,スプライシング阻害剤の処理により,転写伸長に必要なRNAポリメラーゼIIのリン酸化が低下し,転写伸長が抑制されることなども報告されており,スプライシングと転写伸長の間には密接な関連があることが知られている38–40).サイクリンEやE2F1も,いずれかの機構により転写伸長が抑制されていると考えられる.このように,スプライシング阻害により成熟型のmRNAが産生されないだけでなく,それより上流の転写の段階での制御により遺伝子発現が低下することも明らかとなりつつあり,スプライシングが遺伝子発現に与える影響は当初の想定よりもはるかに多岐にわたるといえる.
また,最近の研究から,スプライシングと遺伝子発現機構の新たな関連も明らかとなった.以下にその一例を紹介する.上述のように,スプライシング阻害によるp27*の発現がG1期停止の一因であると報告されたが,同時に全長のp27自体の発現も上昇することも報告された.このp27の発現上昇はどのようになされているのだろうか? タンパク質の量が増加するためには,タンパク質の産生が増加するか,分解が抑制されるかのいずれかだと考えられる.p27は,ユビキチン–プロテアソーム系によりそのタンパク質量が調節されており,また,そのユビキチン化には,CDK2-サイクリンE複合体によるリン酸化が重要である41).上述のように,スプライシング阻害細胞中ではサイクリンEの発現が低下していることから,p27の分解が抑制された結果,p27のタンパク量が増加すると予想された.実際に,スプライシング阻害細胞中では,p27はわずかに安定化していたものの,p27の発現上昇をすべて説明できるほどの顕著な安定化ではなかった42).一方,SSA処理によりp27をコードするCDKN1B mRNAの量が顕著に増加することが明らかとなった.SSAは,当初転写を活性化する化合物として単離され,その後スプライシング阻害剤であることが明らかとなった経緯から,CDKN1B遺伝子の転写が活性化されることによりmRNA量が増加すると思われた9).しかしながら,予想とは異なり,レポーター遺伝子を用いた実験などから,スプライシング阻害はCDKN1B遺伝子の転写には影響を与えないことが明らかとなった.それでは,CDKN1B mRNAの安定性がスプライシング阻害により変化するのであろうか? CDKN1B mRNAの安定性を検証した結果,CDKN1B mRNAは非常に安定性が低く半減期の短いmRNAであることや,スプライシング阻害により安定性が増すことが明らかとなった42).さらに,CDKN1B mRNAの安定性は,その5′UTRや3′UTRにより制御されていることが報告されているが,スプライシング阻害による安定化には,3′UTRの比較的上流領域が関与していることも明らかとなった42–44)(図3).この領域には,CDKN1B遺伝子発現を制御することが報告されているmiR-221とmiR-222が認識する配列があり,これらのmiRNAがスプライシング阻害時のCDKN1B mRNAの安定化に関与している可能性が考えられる45–47).たとえば,miR-221とmiR-222の量が低下する,もしくはmiRNAが働くのに必須なAGO1やGW182などの因子が減少するということが考えられる.miR-221とmiR-222は,さまざまながん細胞での高発現が観察され,p27をはじめとした多くのがん抑制因子の発現を低下させることにより,がんを促進する方向に働くoncomirの一つである48).これらのmiRNAの関与は今後の研究により明らかになると期待されるが,もし実際にスプライシング阻害によりこれらのmiRNAが減少するのであれば,スプライシング阻害剤による抗がん活性の分子メカニズムが非常に幅広いものであることが明らかになるであろう.
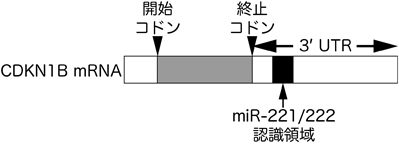
SSAはCDKN1B mRNAの3′UTRを介してCDKN1B mRNAを安定化させる.安定化に必要な領域にはmiR-221/222が認識する領域が存在する.
上述のように,p27*はスプライシング阻害によるG1期停止に関与することが明らかとなっているが,G2/M期停止には関与するのであろうか? 上述のように,p27はG1期からS期への移行をコントロールすることがよく知られているが,興味深いことに,p27*はG1期のみならずG2/M期の停止にも関わっていることが最近の研究から明らかとなった.
細胞周期を同調していない細胞をスプライシング阻害剤処理した結果,細胞がG1期およびG2/M期で停止することが明らかになっていたものの,詳細な解析は行われていなかった5).細胞周期を同調した細胞を用いた研究の結果,G2/M期の細胞にスプライシング阻害剤処理を行うと細胞は次のG1期で停止し,G1期からS期にかけての細胞を処理すると,直後のG2/M期で停止することが明らかとなった36, 49).また,面白いことにスプライシング阻害剤の濃度が比較的高いとG2期で停止し,比較的薄いとM期で停止することも明らかとなり,濃度依存的な細胞周期停止のメカニズムがあることが示唆される.さらに,G2/M期停止に関わる細胞周期関連因子を探索したところ,G2期で停止した細胞においてp27*が蓄積していることが明らかとなった.このp27*の過剰発現はG1期停止同様にG2期停止も引き起こすものの,p27*のノックダウンはスプライシング阻害によるG2/M期停止を解除することはできなかった.このことから,p27*はG2/M期で細胞周期を停止させる能力はあるが,p27*の発現だけがG2/M期停止の原因ではないと考えられる.それでは,p27*はG2/M期停止に関わっているのだろうか? この証明のためには,少々トリッキーな実験が必要であった.SSAとPla-Bは,いずれもその細胞内標的分子であるU2 snRNPに結合しその機能を阻害することでスプライシングを阻害するが,大きな違いが一つある.それは,SSAは標的タンパク質に共有結合するが,Pla-Bは非共有結合であるという点である50).したがって,Pla-Bで細胞を処理したのち培地からPla-Bを除くことで,スプライシング活性は回復する.一方,p27*は非常に安定なタンパク質であるので,Pla-Bを除いてもしばらくの間は細胞内に存在する.その時間差を利用して,スプライシングは正常であるが,p27*は細胞内に存在するという状況を作り出すことができる.その際に,p27*をsiRNAでノックダウンしてしまえば,p27*の効果を評価することができる.少々手の込んだ実験であるが,この方法を用いることにより,p27*の発現がG2/M期停止に関わっていることが確認された.もちろん,上述のようにp27*の発現だけがG2/M期停止の原因ではないので,今後の研究によりさらなるメカニズムの解明が待たれるところである.
上述のように,p27*はp27同様,CDK2-サイクリンEのリン酸化活性を阻害することができる.この阻害活性が,p27*によるG1期停止の分子メカニズムであると考えられるが,G2期停止に関してはどうなのだろうか? G2期からM期への進行にはCDK1-サイクリンAやCDK1-サイクリンBの活性が必要である.最近の研究から,p27*はこれらのCDK複合体に結合し,そのリン酸化活性を阻害することが明らかとなった49).以前の研究で,p27のユビキチン化とプロテアソームによる分解に必要なユビキチンリガーゼの欠損により,p27がG2期において蓄積しCDK複合体を阻害することで,細胞周期が停止することが報告されている51).したがって,今回のケースもp27*がこれらCDK複合体を阻害することがG2期停止を引き起こしていることが強く示唆される.
次に,p27とp27*の違いについて話を進めていこう.上述のように,p27とp27*はいずれもCDK2-サイクリンE, CDK1-サイクリンAやCDK1-サイクリンBを阻害できるので,CDKインヒビターとしての機能的な違いはないと考えられる(図4).それでは,p27とp27*の違いはどこにあるのだろうか? 上でも述べたように,CDKインヒビターとしての機能に重要なN末端側は共通であるが,p27のC末端側はエクソン2に由来する配列であり,p27*のC末端側はイントロン1に由来する配列である5).このC末端がp27とp27*のタンパク質としての安定性に大きな影響を与えているようである.p27のC末端にはリン酸化サイトがあり,このサイトのリン酸化が引き金となり,p27はユビキチン化を受け,プロテアソームにより分解される41)(図2).しかしながら,このサイトはエクソン2に由来する配列中にあり,p27*には存在しないものである.したがって,p27*はp27と比較すると非常に安定だと想定される.実際に,プロテアソーム阻害剤のMG132で細胞を処理することにより,p27のタンパク質量は増加するものの,p27*の量は増加しない49).このことは,p27はプロテアソームの基質であるが,p27*はそうではないことを示している.さらに,ユビキチン化されたp27とp27*の量を比較すると,明らかにユビキチン化p27*の方が少ないということもわかっている.これらのことから,p27*はユビキチン-プロテアソーム系によるタンパク質分解を免れることにより,全長のp27が存在しないG2期にも蓄積することができ,その結果,G2期停止を引き起こしていると考えられる(図4).
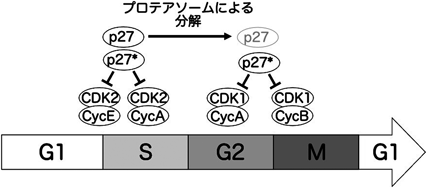
p27*はCDK1-サイクリンA/Bを阻害してG2期停止を引き起こす.p27*はプロテアソームによる分解を免れるため,G2期でも細胞内に存在しCDKを阻害できる.
ここまで述べてきたように,さまざまな生物種において,スプライシングと細胞周期進行には非常に密接な関係があることが示されてきた.しかしながら,スプライシングはほぼすべての遺伝子の発現に影響を与えるのに,細胞周期停止という特定の表現型が観察されるのはなぜなのだろうか? 筆者は,タンパク質の半減期にその原因があるのではないかと考えている.もし,タンパク質の半減期が長ければ,スプライシングが阻害されていたとしても,タンパク質量が減少するまでにはかなりの時間がかかる.一方,細胞周期関連因子の多くは,周期的な発現パターンを示しており,サイクリンなどのように役目が終わると積極的に分解されるものも多くある.したがって,そのような半減期の短いタンパク質は,スプライシング異常により新規タンパク質の合成が阻害されると,急激にタンパク質量が減少してしまうと考えられる.その結果,細胞周期停止が目立った表現型として観察されるのではないだろうか?
また,p27*が生成されることも原因の一つだと考えられる.通常,イントロン部分が翻訳されたタンパク質は機能を持たないと考えられるが,p27*は全長のp27と共通の部分がほとんどであり,生理的な機能を持つ非常に安定なタンパク質である.言い換えれば,p27*は非常に特殊なトランケート型タンパク質なのである.今後,細胞周期進行のみならず,他の細胞機能に影響を与えるトランケート型タンパク質も発見されるかもしれない.今後の研究を通してスプライシングを新たな角度から見直すことにより,面白い発見がなされるのではないかと期待している.
1) Grzybowska, E.A. (2012) Human intronless genes: functional groups, associated diseases, evolution, and mRNA processing in absence of splicing. Biochem. Biophys. Res. Commun., 424, 1–6.
2) Papasaikas, P. & Valcarcel, J. (2016) The spliceosome: The ultimate RNA chaperone and sculptor. Trends Biochem. Sci., 41, 33–45.
3) Wahl, M.C., Will, C.L., & Luhrmann, R. (2009) The spliceosome: Design principles of a dynamic RNP machine. Cell, 136, 701–718.
4) Petasny, M., Bentata, M., Pawellek, A., Baker, M., Kay, G., & Salton, M. (2021) Splicing to keep cycling: The importance of pre-mRNA splicing during the cell cycle. Trends Genet., 37, 266–278.
5) Kaida, D., Motoyoshi, H., Tashiro, E., Nojima, T., Hagiwara, M., Ishigami, K., Watanabe, H., Kitahara, T., Yoshida, T., Nakajima, H., et al. (2007) Spliceostatin A targets SF3b and inhibits both splicing and nuclear retention of pre-mRNA. Nat. Chem. Biol., 3, 576–583.
6) Kotake, Y., Sagane, K., Owa, T., Mimori-Kiyosue, Y., Shimizu, H., Uesugi, M., Ishihama, Y., Iwata, M., & Mizui, Y. (2007) Splicing factor SF3b as a target of the antitumor natural product pladienolide. Nat. Chem. Biol., 3, 570–575.
7) Mizui, Y., Sakai, T., Iwata, M., Uenaka, T., Okamoto, K., Shimizu, H., Yamori, T., Yoshimatsu, K., & Asada, M. (2004) Pladienolides, new substances from culture of Streptomyces platensis Mer-11107. III. In vitro and in vivo antitumor activities. J. Antibiot. (Tokyo), 57, 188–196.
8) Nakajima, H., Hori, Y., Terano, H., Okuhara, M., Manda, T., Matsumoto, S., & Shimomura, K. (1996) New antitumor substances, FR901463, FR901464 and FR901465. II. Activities against experimental tumors in mice and mechanism of action. J. Antibiot. (Tokyo), 49, 1204–1211.
9) Nakajima, H., Sato, B., Fujita, T., Takase, S., Terano, H., & Okuhara, M. (1996) New antitumor substances, FR901463, FR901464 and FR901465. I. Taxonomy, fermentation, isolation, physico-chemical properties and biological activities. J. Antibiot. (Tokyo), 49, 1196–1203.
10) Schneider-Poetsch, T., Chhipi-Shrestha, J.K., & Yoshida, M. (2021) Splicing modulators: on the way from nature to clinic. J. Antibiot. (Tokyo), 74, 603–616.
11) Seiler, M., Yoshimi, A., Darman, R., Chan, B., Keaney, G., Thomas, M., Agrawal, A.A., Caleb, B., Csibi, A., Sean, E., et al. (2018) H3B-8800, an orally available small-molecule splicing modulator, induces lethality in spliceosome-mutant cancers. Nat. Med., 24, 497–504.
12) Hochegger, H., Takeda, S., & Hunt, T. (2008) Cyclin-dependent kinases and cell-cycle transitions: Does one fit all? Nat. Rev. Mol. Cell Biol., 9, 910–916.
13) Lim, S. & Kaldis, P. (2013) Cdks, cyclins and CKIs: Roles beyond cell cycle regulation. Development, 140, 3079–3093.
14) Neumann, B., Walter, T., Heriche, J.K., Bulkescher, J., Erfle, H., Conrad, C., Rogers, P., Poser, I., Held, M., Liebel, U., et al. (2010) Phenotypic profiling of the human genome by time-lapse microscopy reveals cell division genes. Nature, 464, 721–727.
15) Takahashi, K., Yamada, H., & Yanagida, M. (1994) Fission yeast minichromosome loss mutants mis cause lethal aneuploidy and replication abnormality. Mol. Biol. Cell, 5, 1145–1158.
16) Shea, J.E., Toyn, J.H., & Johnston, L.H. (1994) The budding yeast U5 snRNP Prp8 is a highly conserved protein which links RNA splicing with cell cycle progression. Nucleic Acids Res., 22, 5555–5564.
17) Potashkin, J., Kim, D., Fons, M., Humphrey, T., & Frendewey, D. (1998) Cell-division-cycle defects associated with fission yeast pre-mRNA splicing mutants. Curr. Genet., 34, 153–163.
18) Lundgren, K., Allan, S., Urushiyama, S., Tani, T., Ohshima, Y., Frendewey, D., & Beach, D. (1996) A connection between pre-mRNA splicing and the cell cycle in fission yeast: cdc28+ is allelic with prp8+ and encodes an RNA-dependent ATPase/helicase. Mol. Biol. Cell, 7, 1083–1094.
19) Boger-Nadjar, E., Vaisman, N., Ben-Yehuda, S., Kassir, Y., & Kupiec, M. (1998) Efficient initiation of S-phase in yeast requires Cdc40p, a protein involved in pre-mRNA splicing. Mol. Gen. Genet., 260, 232–241.
20) Dominguez, D., Tsai, Y.H., Weatheritt, R., Wang, Y., Blencowe, B.J., & Wang, Z. (2016) An extensive program of periodic alternative splicing linked to cell cycle progression. eLife, 5, e10288.
21) Karni, R., de Stanchina, E., Lowe, S.W., Sinha, R., Mu, D., & Krainer, A.R. (2007) The gene encoding the splicing factor SF2/ASF is a proto-oncogene. Nat. Struct. Mol. Biol., 14, 185–193.
22) Li, X., Wang, J., & Manley, J.L. (2005) Loss of splicing factor ASF/SF2 induces G2 cell cycle arrest and apoptosis, but inhibits internucleosomal DNA fragmentation. Genes Dev., 19, 2705–2714.
23) Colwill, K., Pawson, T., Andrews, B., Prasad, J., Manley, J.L., Bell, J.C., & Duncan, P.I. (1996) The Clk/Sty protein kinase phosphorylates SR splicing factors and regulates their intranuclear distribution. EMBO J., 15, 265–275.
24) Xu, L., Zhang, H., Mei, M., Du, C., Huang, X., Li, J., Wang, Y., Bao, S., & Zheng, H. (2018) Phosphorylation of serine/arginine-rich splicing factor 1 at tyrosine 19 promotes cell proliferation in pediatric acute lymphoblastic leukemia. Cancer Sci., 109, 3805–3815.
25) Chanarat, S. & Strasser, K. (2013) Splicing and beyond: the many faces of the Prp19 complex. Biochim. Biophys. Acta, 1833, 2126–2134.
26) Tsai, W.Y., Chow, Y.T., Chen, H.R., Huang, K.T., Hong, R.I., Jan, S.P., Kuo, N.Y., Tsao, T.Y., Chen, C.H., & Cheng, S.C. (1999) Cef1p is a component of the Prp19p-associated complex and essential for pre-mRNA splicing. J. Biol. Chem., 274, 9455–9462.
27) McDonald, W.H., Ohi, R., Smelkova, N., Frendewey, D., & Gould, K.L. (1999) Myb-related fission yeast cdc5p is a component of a 40S snRNP-containing complex and is essential for pre-mRNA splicing. Mol. Cell. Biol., 19, 5352–5362.
28) Mu, R., Wang, Y.B., Wu, M., Yang, Y., Song, W., Li, T., Zhang, W.N., Tan, B., Li, A.L., Wang, N., et al. (2014) Depletion of pre-mRNA splicing factor Cdc5L inhibits mitotic progression and triggers mitotic catastrophe. Cell Death Dis., 5, e1151.
29) Huang, R., Xue, R., Qu, D., Yin, J., & Shen, X.Z. (2017) Prp19 arrests cell cycle via Cdc5L in hepatocellular carcinoma cells. Int. J. Mol. Sci., 18, 778.
30) Bernstein, H.S. & Coughlin, S.R. (1997) Pombe Cdc5-related protein. A putative human transcription factor implicated in mitogen-activated signaling. J. Biol. Chem., 272, 5833–5837.
31) Graub, R., Lancero, H., Pedersen, A., Chu, M., Padmanabhan, K., Xu, X.Q., Spitz, P., Chalkley, R., Burlingame, A.L., Stokoe, D., et al. (2008) Cell cycle-dependent phosphorylation of human CDC5 regulates RNA processing. Cell Cycle, 7, 1795–1803.
32) Boudrez, A., Beullens, M., Groenen, P., Van Eynde, A., Vulsteke, V., Jagiello, I., Murray, M., Krainer, A.R., Stalmans, W., & Bollen, M. (2000) NIPP1-mediated interaction of protein phosphatase-1 with CDC5L, a regulator of pre-mRNA splicing and mitotic entry. J. Biol. Chem., 275, 25411–25417.
33) Beullens, M. & Bollen, M. (2002) The protein phosphatase-1 regulator NIPP1 is also a splicing factor involved in a late step of spliceosome assembly. J. Biol. Chem., 277, 19855–19860.
34) Burns, C.G., Ohi, R., Mehta, S., O’Toole, E.T., Winey, M., Clark, T.A., Sugnet, C.W., Ares, M. Jr., & Gould, K.L. (2002) Removal of a single alpha-tubulin gene intron suppresses cell cycle arrest phenotypes of splicing factor mutations in Saccharomyces cerevisiae. Mol. Cell. Biol., 22, 801–815.
35) Sakai, Y., Tsujita, T., Akiyama, T., Yoshida, T., Mizukami, T., Akinaga, S., Horinouchi, S., Yoshida, M., & Yoshida, T. (2002) GEX1 compounds, novel antitumor antibiotics related to herboxidiene, produced by Streptomyces sp. II. The effects on cell cycle progression and gene expression. J. Antibiot. (Tokyo), 55, 863–872.
36) Satoh, T. & Kaida, D. (2016) Upregulation of p27 cyclin-dependent kinase inhibitor and a C-terminus truncated form of p27 contributes to G1 phase arrest. Sci. Rep., 6, 27829.
37) Kikuchi, K. & Kaida, D. (2021) CCNE1 and E2F1 partially suppress G1 phase arrest caused by spliceostatin a treatment. Int. J. Mol. Sci., 22, 11623.
38) Koga, M., Satoh, T., Takasaki, I., Kawamura, Y., Yoshida, M., & Kaida, D. (2014) U2 snRNP is required for expression of the 3′ end of genes. PLoS One, 9, e98015.
39) Koga, M., Hayashi, M., & Kaida, D. (2015) Splicing inhibition decreases phosphorylation level of Ser2 in Pol II CTD. Nucleic Acids Res., 43, 8258–8267.
40) Kaida, D., Berg, M.G., Younis, I., Kasim, M., Singh, L.N., Wan, L., & Dreyfuss, G. (2010) U1 snRNP protects pre-mRNAs from premature cleavage and polyadenylation. Nature, 468, 664–668.
41) Montagnoli, A., Fiore, F., Eytan, E., Carrano, A.C., Draetta, G.F., Hershko, A., & Pagano, M. (1999) Ubiquitination of p27 is regulated by Cdk-dependent phosphorylation and trimeric complex formation. Genes Dev., 13, 1181–1189.
42) Kaida, D. & Shida, K. (2022) Spliceostatin a stabilizes CDKN1B mRNA through the 3′UTR. Biochem. Biophys. Res. Commun., 608, 39–44.
43) Shi, H., Li, H., Yuan, R., Guan, W., Zhang, X., Zhang, S., Zhang, W., Tong, F., Li, L., Song, Z., et al. (2018) PCBP1 depletion promotes tumorigenesis through attenuation of p27(Kip1) mRNA stability and translation. J. Exp. Clin. Cancer Res., 37, 187.
44) Ziegeler, G., Ming, J., Koseki, J.C., Sevinc, S., Chen, T., Ergun, S., Qin, X., & Aktas, B.H. (2010) Embryonic lethal abnormal vision-like HuR-dependent mRNA stability regulates post-transcriptional expression of cyclin-dependent kinase inhibitor p27Kip1. J. Biol. Chem., 285, 15408–15419.
45) Galardi, S., Mercatelli, N., Giorda, E., Massalini, S., Frajese, G.V., Ciafre, S.A., & Farace, M.G. (2007) miR-221 and miR-222 expression affects the proliferation potential of human prostate carcinoma cell lines by targeting p27Kip1. J. Biol. Chem., 282, 23716–23724.
46) le Sage, C., Nagel, R., Egan, D.A., Schrier, M., Mesman, E., Mangiola, A., Anile, C., Maira, G., Mercatelli, N., Ciafre, S.A., et al. (2007) Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J., 26, 3699–3708.
47) Visone, R., Russo, L., Pallante, P., De Martino, I., Ferraro, A., Leone, V., Borbone, E., Petrocca, F., Alder, H., Croce, C.M., et al. (2007) MicroRNAs (miR)-221 and miR-222, both overexpressed in human thyroid papillary carcinomas, regulate p27Kip1 protein levels and cell cycle. Endocr. Relat. Cancer, 14, 791–798.
48) Garofalo, M., Quintavalle, C., Romano, G., Croce, C.M., & Condorelli, G. (2012) miR221/222 in cancer: their role in tumor progression and response to therapy. Curr. Mol. Med., 12, 27–33.
49) Kaida, D., Satoh, T., Ishida, K., Yoshimoto, R., & Komori, K. (2022) A truncated form of the p27 CDK inhibitor translated from pre-mRNA causes G2-phase arrest. Mol. Cell. Biol., 42, e00217-22.
50) Cretu, C., Gee, P., Liu, X., Agrawal, A., Nguyen, T.V., Ghosh, A.K., Cook, A., Jurica, M., Larsen, N.A., & Pena, V. (2021) Structural basis of intron selection by U2 snRNP in the presence of covalent inhibitors. Nat. Commun., 12, 4491.
51) Nakayama, K., Nagahama, H., Minamishima, Y.A., Miyake, S., Ishida, N., Hatakeyama, S., Kitagawa, M., Iemura, S., Natsume, T., & Nakayama, K.I. (2004) Skp2-mediated degradation of p27 regulates progression into mitosis. Dev. Cell, 6, 661–672.

富山大学学術研究部医学系 准教授.博士(理学).
1998年東京大学理学部生物学科卒業.2003年同大学院理学系研究科生物科学専攻修了,理化学研究所博士研究員.07年ペンシルベニア大学博士研究員.11年富山大学テニュアトラック助教.16年より現職.
研究テーマと抱負スプライシング阻害剤などを用いたケミカルバイオロジー研究を行なっている.化合物を用いることで,通常の手法では明らかにできなかった生命の仕組みを一つでも多く明らかにしていきたい.
ウェブサイトhttp://www.kaida-lab.com
趣味マラソン,子供と遊ぶこと.

This page was created on 2022-11-18T15:46:46.733+09:00
This page was last modified on 2022-12-14T11:25:00.000+09:00
このサイトは(株)国際文献社によって運用されています。