窒素栄養に応答した植物の花成制御Flowering regulation in response to nitrogen availability
北海道大学大学院理学研究院Faculty of Science, Hokkaido University ◇ 〒060–0810 札幌市北区北10条西8丁目 ◇ Kita-10 Nishi-8, Kita-ku, Sapporo 060–0810, Japan
発行日:2022年12月25日Published: December 25, 2022
© 2022 公益社団法人日本生化学会© 2022 The Japanese Biochemical Society
窒素は,植物にとって最も要求量の多い栄養素であり,欠乏すると生育や作物収量に大きな影響を与える.そのため,農業では施肥による窒素の供給が行われている.しかし,窒素を過剰に与えると,葉を大きく成長させる一方で花つきが悪くなり収量が低下することが知られており,農業で古くから問題となっている.そのため,適切な時期に花成を誘導することは,作物収量を最適化するために施肥管理において重要な点となっている.また,窒素応答性花成は,細胞内の栄養状態がどのようにして植物の発生制御に作用するのかという問いとして,植物生理学的にも重要な未解決課題である.本稿では,窒素量に応じた花成制御の分子機構に関して,筆者らの研究内容を含めた最近の知見を紹介する.
花成とは,栄養成長相から生殖成長相への移行であり,植物の発生において重要な転換点である.シロイヌナズナにおいて,花成は温度,光周期,加齢,ジベレリンといったさまざまなシグナルによって制御されていることがわかっている1).しかし,いずれのシグナル伝達経路も,葉でのFT(FLOWERING LOCUS T)の増加または茎頂メリステムでのSOC1(SUPRESSOR OF OVEREXPRESSION OF CONSTANS 1)の増加に収束することがわかっている2).FTは,花成ホルモン「フロリゲン」の実体で,葉の維管束伴細胞で発現したあと茎頂メリステムに輸送され,花芽形成に必要な遺伝子発現を誘導する3).
農業現場では,窒素施肥が原因で葉が成長過多となり,生殖成長が遅れて作物収量が低下することが知られており,施肥管理において重要となっている.つまり,窒素は花成のブレーキ役を担い,窒素が少ないときには花成が早期化するという関係性である.その一方で,シロイヌナズナをはじめ,これまでに窒素と花成時期の関係を調べた研究の結果としては,低窒素条件で花成が遅れる現象も報告されており,統一的な見解がなされていなかった.近年,こうした窒素量に応じた花成時期の変化に関して,「U字型花成応答」と呼ばれるモデルが提唱された4).植物は,ある一定量以上の窒素獲得量の範囲では,高窒素よりも低窒素条件下で花成が早期化するが,一定量以下になると逆に花成が遅延するというものである.このU字型花成応答を引き起こす窒素濃度は,窒素源や植物の栽培方法,植物種によっても異なると考えられている.U字型花成応答の制御機構の全容はまだ不明であるが,モデル植物での遺伝学的解析や生化学的解析から,少しずつそれに関与する分子機能がわかってきている.
Yuanらは,窒素条件に応じたトランスクリプトーム解析から,窒素応答性花成に関与する因子としてシロイヌナズナのフェレドキシン-NADP+還元酵素FNR1(FERREDOXIN-NADP+-OXIDOREDUCTASE)と青色光受容体CRY1(Cryptochrome 1)を同定した5).低窒素条件では,FNR1およびCRY1の発現量が増加し,加えて核内CRY1タンパク質量が増加することで,下流に位置する概日時計遺伝子の発現パターンが変化し,最終的に花成が促進されるというモデルが提唱されている.
また,窒素による花成促進について興味深い報告がある.Olasらは,窒素が不足しすぎると,茎頂メリステムにおけるSOC1の発現が減少することを見いだした6).そして,窒素に応じたSOC1の発現変動は,硝酸シグナル伝達のマスターレギュレーターである転写因子NLP7(NIN-LIKE PROTEIN 7)/NLP6の制御下にあることが示された.つまり,窒素量は,硝酸シグナルを介して,茎頂メリステムにおける花芽形成遺伝子の発現を正に制御することが示された.こうした制御が,窒素が不足しすぎた際の花成遅延を引き起こす原因の一つと考えられる.プロモーター領域の配列から,SOC1の上流転写因子SPL(SQUAMOSA PROMOTER BINDING-LIKE)3/SPL5の発現がNLPによる直接的な制御を受ける可能性が示唆されているが,詳しい関係はわかっていない.
このように,葉や茎頂メリステムにおける花成関連因子が,それぞれ異なる経路を介して窒素量に応答し,そうした総和によって,窒素量に応じた花成時期が決定されることが予想される.
また,最近イネにおいても窒素応答性花成制御に関わる転写因子としてNhd1(N-mediated heading date 1)が報告された7).Nhd1は,グルタミンによって遺伝子発現が上昇し,イネのFTホモログであるHd3a(Heading date 3a)遺伝子8)の転写を直接的に活性化することで花成を誘導することがわかった.ただし,Nhd1の知見だけでは,窒素量によるイネ花成遅延等の表現型と一致しない部分もあり,イネのU字型花成応答を説明するには至っていない.
このように,窒素応答性花成制御に関わる因子の報告はいくつかあるものの,依然としてその全容は不明である.特に,窒素による花成のブレーキ機構(=低窒素による花成促進)に関する詳細は謎が多く残っている.
筆者らは,シロイヌナズナを用いて,低窒素による花成誘導に着目し,研究を行った.まず,細胞内のタンパク質リン酸化状態を網羅的に検出するリン酸化プロテオーム解析データ9)から,窒素と花成をつなぐシグナル伝達因子を探索した.その結果,低窒素下でリン酸化状態が変動するタンパク質として転写因子FBH4(FLOWERING BHLH 4)を同定した.FBHはbHLH型転写因子で,シロイヌナズナのゲノムには少なくともFBH1, FBH2, FBH3, FBH4の四つのFBH遺伝子がコードされている.これらFBHsは,CO(CONSTANS)遺伝子の直接的な転写活性化因子であり,FBHsが冗長的に機能することが報告されている10).COはFTの転写活性化因子であり,光周期依存的花成ではこのCO-FT経路の活性化が重要となっている11).そこで,まずFBHsの変異株を用いて低窒素応答性花成の表現型を確認した.FBHs四重機能抑制株を用いて窒素に応じた花成時期の評価を行った結果,低窒素条件下において野生型株で花成の早期化が誘導されたのに対し,FBHs四重機能抑制株ではこのような花成誘導が抑制されていた.この際,野生型株では低窒素条件でCOとFT遺伝子発現が上昇しており,特に夕方から夜間の時間帯にかけて顕著な遺伝子発現量の増加がみられた.これは,FBHsがCOの転写を活性化する時間帯と一致する.実際に,FBHs四重機能抑制株では,低窒素でのCOおよびFTの遺伝子発現量増加がみられなかった.さらに,CO機能欠損株で同様の低窒素条件での花成時期の評価を行った結果,CO機能欠損株では低窒素での花成早期化がみられなかった.これらの結果から,低窒素条件でFBHsがCOおよびFTの発現上昇を介して花成を促進することが示唆された.
次に筆者らは,低窒素誘導性花成におけるFBH4のリン酸化修飾の意義について解析を行った.FBH4は,リン酸化プロテオーム解析で同定された部位(Ser26)以外にも多数のセリン・トレオニン残基を含んでおり,Phos-tagを用いたウェスタンブロット解析から複数部位で高度にリン酸化されていることがわかった.また,Ser26をアラニンに置換したFBH4 S26Aでは,COへの転写活性に変化はみられなかった.そこで,シロイヌナズナFBHsおよび他植物種ホモログ内での保存性から重要性が予想された10か所のセリン・トレオニン残基をまとめてアラニンに置換したFBH4 10Aでの解析を行った.その結果,FBH4 10Aでは,Phos-tagウェスタンブロットで検出されるリン酸化バンドが減っており,CO転写活性が顕著に増加していた.つまり,FBH4のリン酸化は,自身のCO転写活性を負に制御していることが示唆された.加えて,細胞内局在性解析からもFBH4リン酸化に関する興味深い結果が得られた.各窒素条件でのFBH4細胞内局在性を解析したところ,通常窒素条件でFBH4は主に細胞質に局在しているが,低窒素条件では核内FBH4量が増加することがわかった.そして,核と細胞質ではFBH4のリン酸化状態が異なり,核ではリン酸化度合いが低いFBH4が多く存在することがわかった.これと一致して,FBH4 10A型は細胞質より核に多く局在していた.これらの結果から,低窒素条件では特定部位のFBH4リン酸化レベルが減少することで,核内のFBH4が増加し,CO転写を活性化することで花成を誘導することが示された.
上記の解析から,低窒素誘導花成にはFBH4のリン酸化状態が分子スイッチとしての役割を果たすことが示唆された.では,その上流で細胞内の窒素量を感知し,FBH4のリン酸化レベルを制御する機構はどのようなものだろうか.これまで,花成を誘導するような長期的な低窒素状態の感受やシグナル伝達の分子機構についてはあまりわかっていない.筆者らは,この点についてさらなる解析を行い,FBH4をリン酸化する上流キナーゼとしてSnRK1(SNF1-RELATED PROTEIN KINASE 1)を同定した.SnRK1は,哺乳類AMPK(AMP-ACTIVATED KINASE)や酵母SNF1(SUCROSE NON-FERMENTING 1)の植物オルソログであり,真核生物に広く保存されたセリン・トレオニンキナーゼである12).AMPK/SNF1/SnRK1は細胞内の代謝恒常性維持に重要な働きを持つエネルギーセンサーであることが知られており,酵素活性を持つαサブユニットと調節作用を持つβおよびγサブユニットからなる三量体を形成して機能する13).ただし,SnRK1を構成するサブユニットのドメイン構造は,AMPKやSNF1とは異なるため,SnRK1の活性がどのようなリガンドによって制御されているかは不明点が多い.実際,SnRK1では,AMPKがリガンドとするAMPによるアロステリックな活性化がみられない14).また,窒素シグナルとSnRK1の関連についてはあまりわかっていなかった.筆者らが行った上述のリン酸化プロテオーム解析より,FBH4に加えて,窒素条件に応じたSnRK1活性変動の可能性が示唆された9).また,Yuanらの研究で,SnRK1が窒素応答性花成制御に関与していることも示唆された5).そこで,低窒素条件下の植物体内におけるSnRK1活性変動について解析した.これまでの研究ではSnRK1活性は,AMPKやSNF1のキナーゼ活性と相関が示されている触媒ドメイン活性化ループ内のリン酸化や下流マーカー遺伝子の発現変化から評価されていた.しかし,植物のSnRK1はAMPKやSNF1と異なり,キナーゼ活性と触媒ドメイン活性化ループ内のリン酸化レベルに必ずしも相関がみられない場合が確認されており,他の活性制御機構の存在も示唆されている.そこで,筆者らはSnRK1活性を直接的に評価できる新たな手法としてACCレポーターを開発した.ACCレポーターでは,AMPK/SNF1/SnRK1によってリン酸化されるラットのACC1(acetyl-CoA carboxylase 1)ペプチドを標的とし,リン酸化特異的ACC抗体でリン酸化レベルを検証することで,SnRK1活性が評価できる.このレポーターを発現するシロイヌナズナ株を作出し,SnRK1活性の解析を行った.その結果,低窒素条件ではSnRK1活性が低下することがわかった.さらに,SnRK1機能抑制株では,野生型に比べてCOおよびFT遺伝子発現が上昇していることがわかった.
では,SnRK1はFBH4のリン酸化変動に関与しているのだろうか.まず,プロトプラスト細胞での一過的発現系においてSnRK1とFBH4の共発現実験を行った結果,SnRK1との共発現により,FBH4によるCO転写活性が顕著に低下した.さらに,SnRK1機能抑制株ではFBH4のリン酸化レベルが低下していることもわかった.そして,シロイヌナズナから精製したSnRK1複合体とリコンビナントFBH4を用いたin vitroでの試験から,SnRK1が直接的にFBH4をリン酸化することもわかった.
以上の結果から,筆者らはSnRK1-FBH4モジュールによる窒素応答性花成制御のモデルを提唱した(図1)15).窒素十分条件では,SnRK1がFBH4をリン酸化することで,FBH4は細胞質に多く存在しており,COへの転写活性は低く維持されている.低窒素条件では,SnRK1の活性が減少することでFBH4のリン酸化レベルが低下し,核局在が亢進し,COとFTの遺伝子発現が促進されることで花成が誘導される.本研究によって,長年不明瞭であった窒素応答性花成制御の分子機構の一端が明らかとなった.
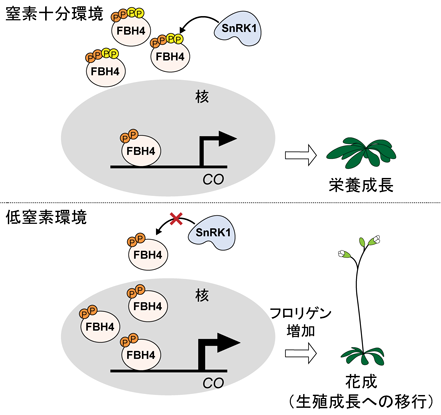
FBH4は,複数部位でリン酸化修飾を受けており,その一部はSnRK1により触媒される.窒素十分環境では,SnRK1がFBH4をリン酸化し,COの転写活性は低く維持されている.低窒素環境では,SnRK1の活性が低下することでFBH4のリン酸化レベルが減少し,FBH4の核局在およびCO転写活性が亢進する.これによりFT(フロリゲン)が増加し,花成が早期化する.P:リン酸化(文献15より改変).
本稿で紹介したSnRK1によるFBH4リン酸化を介した花成制御は,FBH4のリン酸化が窒素過多による花成抑制におけるブレーキ機能を担うことを示唆している.今後,FBH4リン酸化部位とその機能の詳細を明らかにすることで,作物収量改善に向けた新たな施肥最適化技術の開発に貢献したい.また最近筆者らは,FBH4が低窒素条件での花成誘導だけではなく,窒素代謝調節にも関与することを見いだしている.花成時には,成熟葉(ソース器官)から花芽・種子(シンク器官)への栄養素の分配をはじめとした大規模な代謝調節が伴うと予想されるが,花成と代謝を協調的に制御する分子機構は未解明である.今後,花成と代謝におけるFBH4の機能を解析することで,こうした分子機構の解明につながると期待される.
本稿で紹介した筆者らの研究遂行にあたり,共同研究者および北海道大学大学院理学研究院形態機能学講座IIの方々にたくさんのご支援を賜りました.厚く御礼申し上げます.
1) Srikanth, A. & Schmid, M. (2011) Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci., 68, 2013–2037.
2) Amasino, R. (2010) Seasonal and developmental timing of flowering. Plant J., 61, 1001–1013.
3) Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A., Ikeda, Y., Ichinoki, H., Notaguchi, M., Goto, K., & Araki, T. (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science, 309, 1052–1056.
4) Lin, Y.L. & Tsay, Y.F. (2017) Influence of differing nitrate and nitrogen availability on flowering control in Arabidopsis. J. Exp. Bot., 68, 2603–2609.
5) Yuan, S., Zhang, Z.W., Zheng, C., Zhao, Z.Y., Wang, Y., Feng, L.Y., Niu, G., Wang, C.Q., Wang, J.H., Feng, H., et al. (2016) Arabidopsis cryptochrome 1 functions in nitrogen regulation of flowering. Proc. Natl. Acad. Sci. USA, 113, 7661–7666.
6) Olas, J.J., Van Dingenen, J., Abel, C., Działo, M.A., Feil, R., Krapp, A., Schlereth, A., & Wahl, V. (2019) Nitrate acts at the Arabidopsis thaliana shoot apical meristem to regulate flowering time. New Phytol., 223, 814–827.
7) Zhang, S., Zhang, Y., Li, K., Yan, M., Zhang, J., Yu, M., Tang, S., Wang, L., Qu, H., Luo, L., et al. (2021) Nitrogen mediates flowering time and nitrogen use efficiency via floral regulators in rice. Curr. Biol., 31, 671–683.e5.
8) Tamaki, S., Matsuo, S., Hann, L.W., Yokoi, S., & Shimamoto, K. (2007) Hd3a protein is a mobile flowering signal in rice. Science, 316, 1033–1036.
9) Li, X., Sanagi, M., Lu, Y., Nomura, Y., Stolze, S.C., Yasuda, S., Saijo, Y., Schulze, W.X., Feil, R., Stitt, M., et al. (2020) Protein phosphorylation dynamics under carbon/nitrogen-nutrient stress and identification of a cell death-related receptor-like kinase in Arabidopsis. Front. Plant Sci., 11, 377.
10) Ito, S., Song, Y.H., Josephson-Day, A.R., Miller, R.J., Breton, G., Olmstead, R.G., & Imaizumi, T. (2012) FLOWERING BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis. Proc. Natl. Acad. Sci. USA, 109, 3582–3587.
11) Shim, J.S., Kubota, A., & Imaizumi, T. (2017) Circadian clock and photoperiodic flowering in Arabidopsis: CONSTANS is a Hub for Signal integration. Plant Physiol., 173, 5–15.
12) Baena-González, E., Rolland, F., Thevelein, J.M., & Sheen, J. (2007) A central integrator of transcription networks in plant stress and energy signalling. Nature, 448, 938–942.
13) Broeckx, T., Hulsmans, S., & Rolland, F. (2016) The plant energy sensor: evolutionary conservation and divergence of SnRK1 structure, regulation, and function. J. Exp. Bot., 67, 6215–6252.
14) Emanuelle, S., Hossain, M.I., Moller, I.E., Pedersen, H.L., van de Meene, A.M., Doblin, M.S., Koay, A., Oakhill, J.S., Scott, J.W., Willats, W.G., et al. (2015) SnRK1 from Arabidopsis thaliana is an atypical AMPK. Plant J., 82, 183–192.
15) Sanagi, M., Aoyama, S., Kubo, A., Lu, Y., Sato, Y., Ito, S., Abe, M., Mitsuda, N., Ohme-Takagi, M., Kiba, T., et al. (2021) Low nitrogen conditions accelerate flowering by modulating the phosphorylation state of FLOWERING BHLH 4 in Arabidopsis. Proc. Natl. Acad. Sci. USA, 118, e2022942118.

北海道大学大学院理学研究院生物科学部門特任助教.博士(生命科学).
2022年北海道大学大学院生命科学院博士課程修了.同年より現職.
研究テーマと抱負栄養環境に応じた植物の成長相転換制御に関して研究しています.
ウェブサイトhttps://www.sci.hokudai.ac.jp/CSF2-web/
趣味お菓子作り.
北海道大学大学院理学研究院生物科学部門准教授.博士(生命科学).
2010年北海道大学大学院生命科学院博士後期課程修了.同年日本学術振興会特別研究員.11年北海道大学大学院理学研究院生物科学部門助教.20年より現職.
研究テーマと抱負生物がどのように環境情報を伝えて,適応しているのかをタンパク質機能解析から明らかにしたい.
ウェブサイトhttps://www.sci.hokudai.ac.jp/CSF2-web/
趣味音楽鑑賞,散歩.

This page was created on 2022-11-09T13:04:44.393+09:00
This page was last modified on 2022-12-14T10:47:12.000+09:00
このサイトは(株)国際文献社によって運用されています。