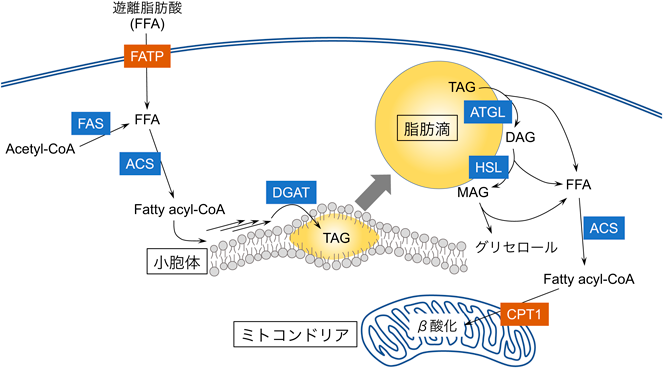
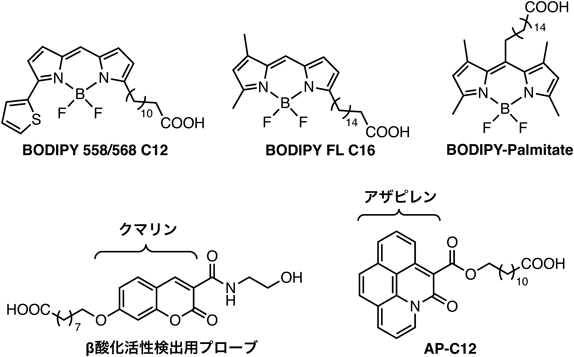
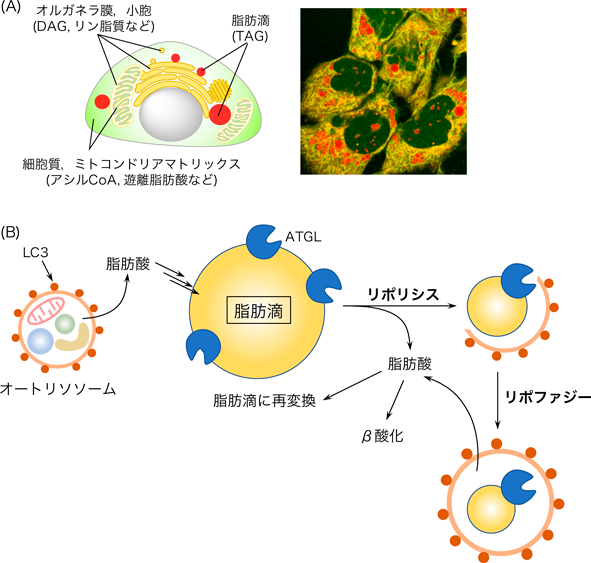
蛍光プローブを用いた脂肪酸代謝解析Fluorescence probes for analyzing fatty acid metabolism
名古屋大学トランスフォーマティブ生命分子研究所Institute of Transformative Bio-Molecules (ITbM), Nagoya University ◇ 〒464–8601 名古屋市千種区不老町 ◇ Furo-cho, Chikusa-ku, Nagoya 464–8601, Japan
発行日:2023年2月25日Published: February 25, 2023