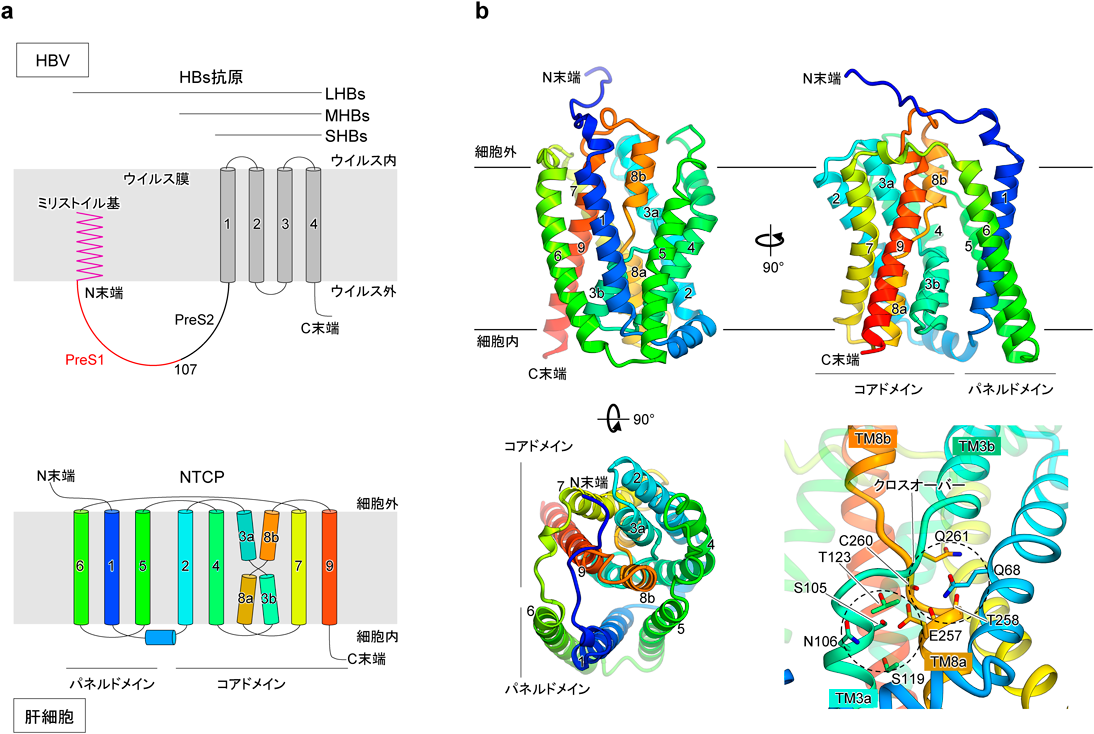
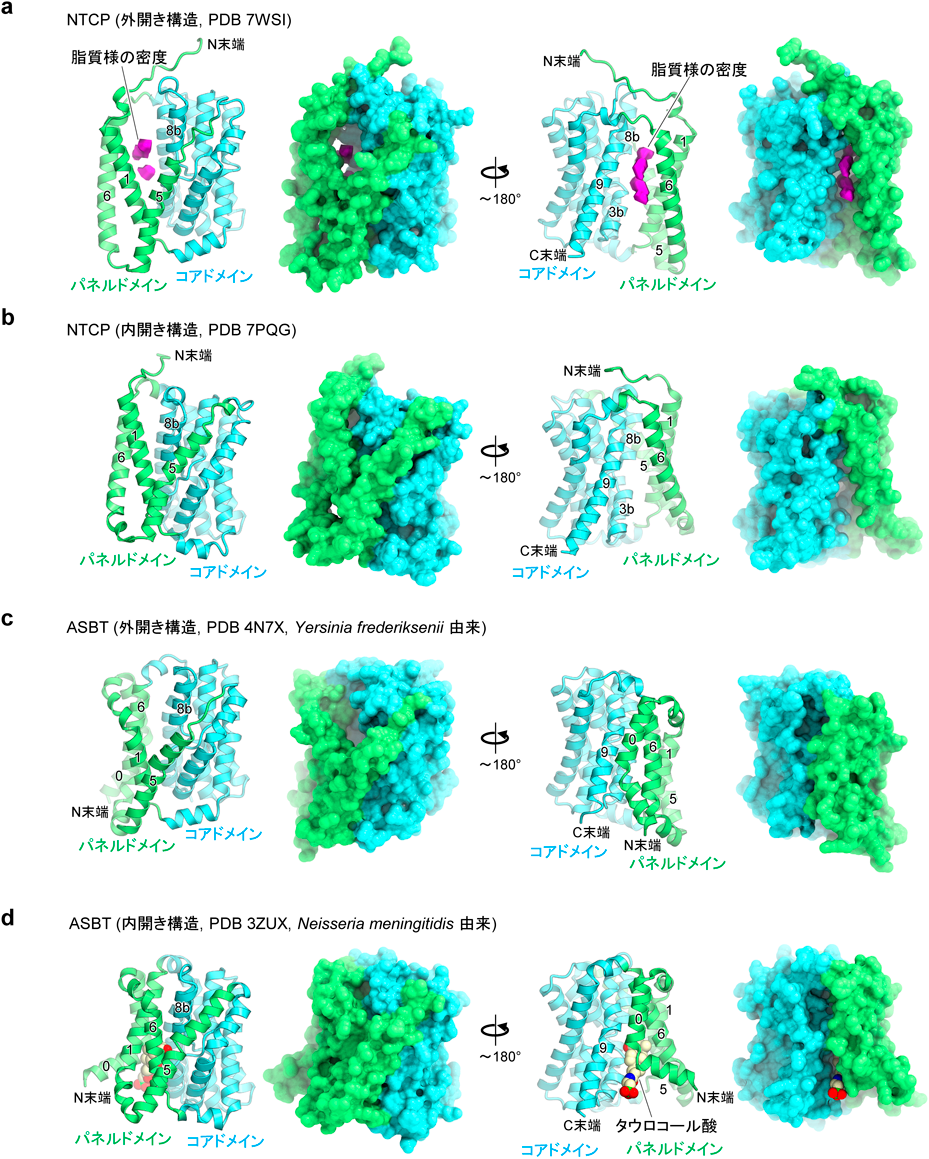
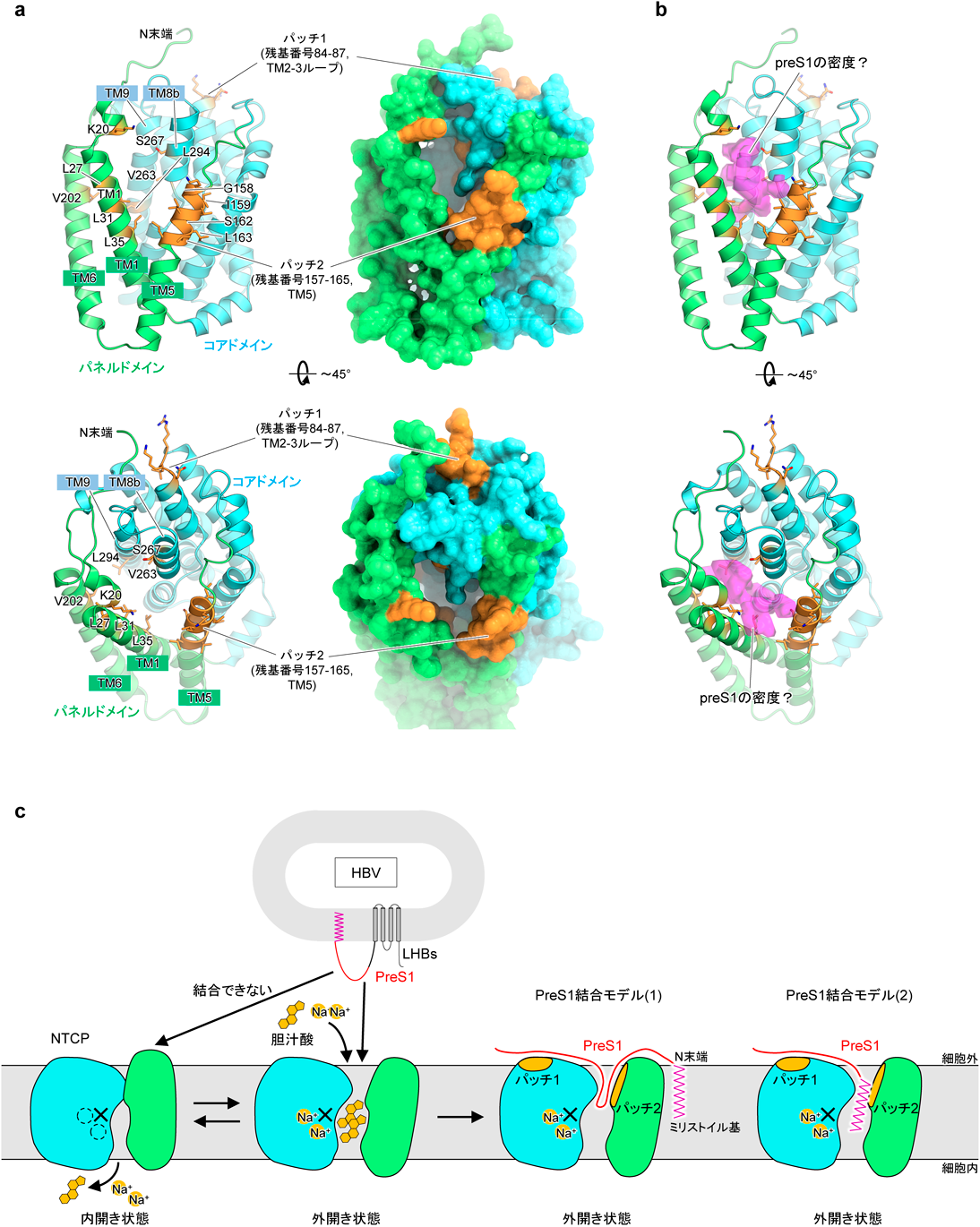
B型肝炎ウイルスの感染受容体NTCPの構造Structure of the hepatitis B virus entry receptor NTCP
東京大学大学院薬学系研究科Graduate School of Pharmaceutical Sciences, The University of Tokyo ◇ 〒113–0033 東京都文京区本郷7–3–1 ◇ 7–3–1 Hongo, Bunkyo-ku, Tokyo 113–0033, Japan
発行日:2023年2月25日Published: February 25, 2023