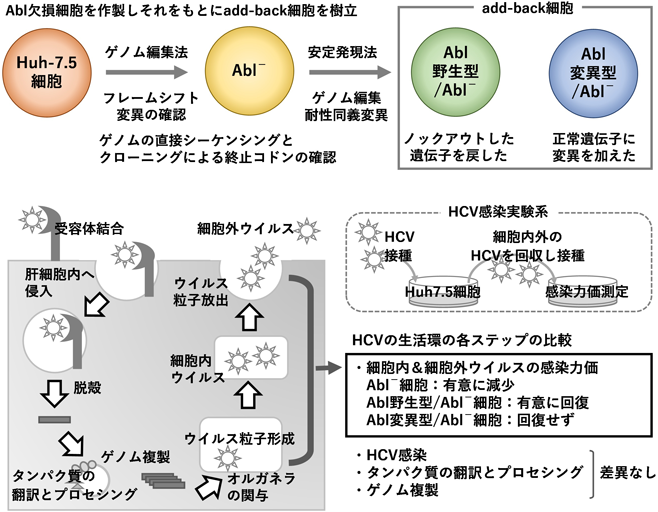
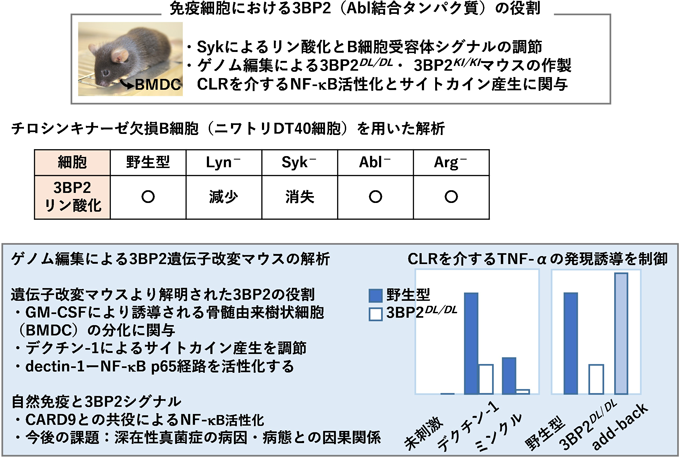
感染宿主因子としてのチロシンキナーゼAblの新しい役割Novel roles of tyrosine kinase Abl as a host factor for microbial infection.
1 福井大学学術研究院医学系部門Department of Genome Science and Microbiology, Faculty of Medical Sciences, University of Fukui ◇ 〒910–1193 福井県吉田郡永平寺町松岡下合月23–3 ◇ 23–3 Matsuoka-Shimoaizuki, Eiheiji, Yoshida-gun, Fukui 910–1193, Japan
2 福井大学ライフサイエンスイノベーション推進機構Organization for Life Science Advancement Programs, University of Fukui ◇ 〒910–1193 福井県吉田郡永平寺町松岡下合月23–3 ◇ 23–3 Matsuoka-Shimoaizuki, Eiheiji, Yoshida-gun, Fukui 910–1193, Japan
発行日:2023年2月25日Published: February 25, 2023