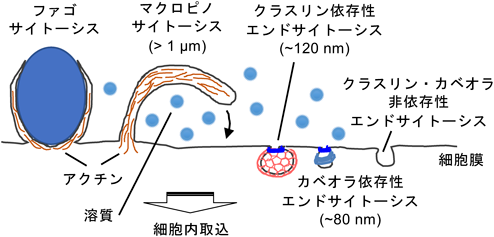
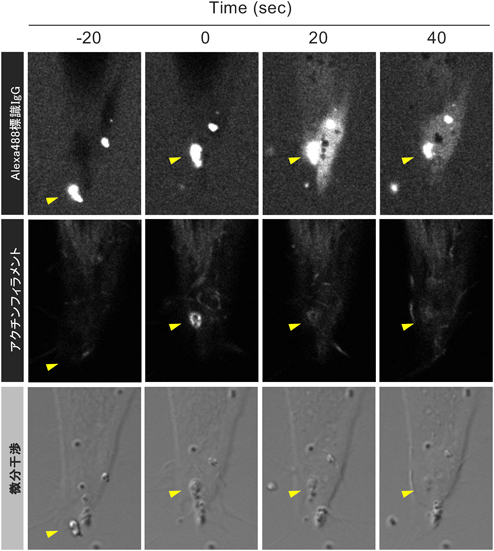
細胞外微粒子の細胞内取り込みとマクロピノサイトーシスContribution of macropinocytosis in the cellular uptake of extracellular fine particles
京都大学化学研究所Institute for Chemical Research, Kyoto University ◇ 〒611–0011 宇治市五ヶ庄 ◇ Uji, Kyoto 611–0011, Japan
発行日:2023年4月25日Published: April 25, 2023