植物発生調節におけるCLEペプチドホルモンの多様な機能Diverse function of CLE peptide hormones in plant growth and development
学習院大学理学部生命科学科Gakushuin University ◇ 〒171–8588 東京都豊島区目白1–5–1 ◇ 1–5–1 Mejiro, Toshima-ku, Tokyo 171–8588, Japan
発行日:2023年4月25日Published: April 25, 2023
© 2023 公益社団法人日本生化学会© 2023 The Japanese Biochemical Society
近年,植物を対象とした生化学・分子遺伝学研究の進展によって,植物は動物とは異なる独自のペプチドホルモンを獲得してきたことがわかってきた.これまでに同定されたペプチドの構造や生理機能は多様であり,また最近新たなペプチドも多数報告されている.植物ペプチドホルモンの分類や定義は現在も発展途上といえるが,一つの定義として「植物の成長や分化,環境応答に関与する内生の分泌型ペプチドのうち,植物界における普遍性の高いもの」1)があげられ,本稿でもこれを踏襲する.
植物ペプチドホルモンはいずれも分泌性の前駆体タンパク質から生じるが,成熟型ホルモンの構造的特徴から「短鎖翻訳後修飾ペプチド」と「システインリッチペプチド」の2種類に大別される1)(図1).前者は,前駆体タンパク質が翻訳後修飾とプロテアーゼによるプロセシングを経て5~20アミノ酸長程度の短いペプチドとして成熟するものである.一方,後者は前駆体タンパク質に多数のシステイン残基が含まれるもので,分子内でのジスルフィド結合形成によって一定の立体構造を保っている.ペプチドホルモンのプロセシングは産生した細胞内で起こる場合が一般的と考えられるが,細胞外プロテアーゼによって切断されて成熟するペプチドホルモンも見つかっている2).植物ペプチドホルモンの多くは産生細胞の近隣の細胞に働きかけるが,一部のペプチドホルモンは維管束を経由して根から地上部へと長距離輸送されることで全身的な情報伝達を担っている3).
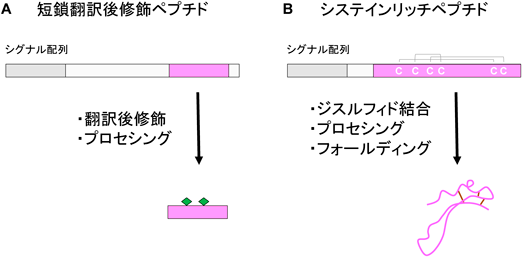
植物ペプチドホルモンの成熟過程の模式図.全長100アミノ酸ほどの前駆体タンパク質は分泌のシグナル配列と将来ホルモンになる領域を含んでいる.(A)短鎖翻訳後修飾ペプチド,(B)システインリッチペプチド.
植物ペプチドホルモンの受容体は主にロイシンリッチリピート型受容体キナーゼ(leucine-rich repeat receptor kinase:LRR-RK)に属している.LRR-RKはI型膜貫通タンパク質であり,細胞外領域のロイシンリッチリピート(LRR)においてペプチドを受容する.植物ゲノムには多数のロイシンリッチリピート型受容体様キナーゼ(LRR receptor-like kinase:LRR-RLK)遺伝子がコードされ,15個のサブクラス(LRR I~XV)に分類される.これまでに報告されたペプチドホルモン受容体はLRR-XおよびLRR-XIに含まれ,いずれも20個ほどのLRRを含む細胞外領域を持つ.これに加え,ペプチドの受容にはLRR-IIサブクラスが共受容体として働くと考えられている.LRR-IIサブクラスは5個程度のLRRからなる比較的短い細胞外領域を持ち,受容体・共受容体間の複合体がリガンド依存的に形成されることによって細胞質側のキナーゼドメインが活性化されるというモデルが提唱されている4).
一般に,生体内ではペプチドホルモンと受容体の分子種が多数共存しているが,特定の受容体は特定のリガンドを認識することで細胞間情報伝達の特異性が生じると考えられる.実際には,特定の植物ペプチドホルモンをコードする遺伝子が植物ゲノム中に複数存在してファミリーを形成していることが一般的である.同一ファミリー内の複数のペプチドは,生体内において冗長的な役割を持つ場合もあるが,生体内での産生部位が異なるために独自の機能に分化している場合も知られている.さらに,植物種間を比較すると,その生理機能が大きく異なっている場合も見つかってきた.筆者は,植物ペプチドホルモンの生理機能が多様化した仕組みを解明することを目的として,ペプチドホルモンの遺伝的多様性に関する研究を行っており,本稿では短鎖翻訳後修飾ペプチドの一群であるCLE(CLAVATA3/EMBRYO SURROUNDING REGION-related)ファミリーに着目した研究について詳述する.
被子植物のモデル生物であるシロイヌナズナにおいて単離されたclavata(clv)変異体群は果実(長角果)がこん棒状に肥大する.1990年代後半に同定された3個のCLV遺伝子座(CLV1, CLV2, CLV3)のうち,CLV1はLRR-RLK, CLV3は約100アミノ酸からなる分泌性タンパク質をコードしていた5, 6).CLV3タンパク質C末端付近の14アミノ酸長の配列と類似の配列を持つタンパク質が,さまざまな植物ゲノム中に多数存在することが判明し,これらはCLEファミリーと名づけられた7).成熟型CLEペプチドは,2000年代後半に複数の日本の研究グループによって報告された.TDIFとCLV3という2個のCLEペプチドが同定され,プロリン水酸化や糖鎖修飾を受けた12~13アミノ酸長のCLEペプチドの成熟型の一次構造が明らかとなった8–10)(図2).
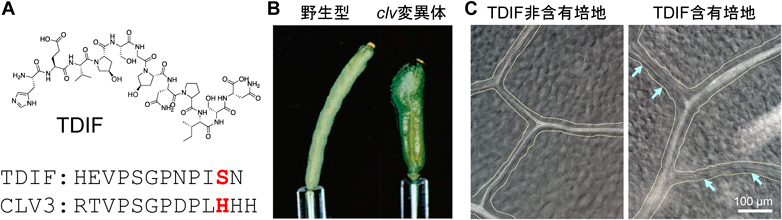
シロイヌナズナにおけるCLEペプチドの構造と生理機能.(A)TDIFの構造とCLV3ペプチドとの比較.TDIFの特異性をもたらす残基の位置を太字で示している.(B)シロイヌナズナclv変異体におけるこん棒状の長角果の表現型.(C)シロイヌナズナの葉脈におけるTDIFの道管細胞分化抑制活性.点線は葉脈を,矢印は道管細胞が形成されていない部分を示す.文献5および12より改変.
TDIF(tracheary element differentiation inhibitory factor)は,維管束中の木部道管細胞の分化を阻害し,幹細胞の維持・増加を促す生理活性を持つ11, 12).一方のCLV3は,地上部のすべての器官を生み出す分裂組織において,幹細胞の細胞分化を促す活性を持ち,結果として花や果実の成長を調節している.TDIFとCLV3はよく似た配列のペプチドであるが,それぞれに特異的なLRR-XIサブクラスの受容体(TDRおよびCLV1)に結合するために,生理機能の特異性が生じている.さらに,LRR-IIサブクラスの共受容体(SERK/CIK)も特異的であり,TDIF–TDR–SERKおよびCLV3–CLV1–CIKというリガンド–受容体–共受容体の組合わせが存在している.TDIF–TDR–SERKの共結晶構造から,TDRのLRRドメインの側面にTDIFが伸展して結合しており,さらにTDIF–TDR複合体をSERKが認識すると考えられている.
CLEペプチドの配列中で生理活性の特異性に重要な役割を担うのが,C末端付近のアミノ酸残基(His11またはSer11)である(図2).TDIFのアラニンスキャン解析において,Ser11はTDIFの生理活性に必要ないことがわかっていた8).しかしながら,このアミノ酸残基は植物種間で高度に保存されており,未知の生物学的な機能が示唆されていた.興味深いことに,Ser11をCLV3型のHis11に置換した人工ペプチドは,TDIFの活性に加えてCLV3と同様に幹細胞分化を促して分裂組織の維持を阻害する活性を示すことが明らかとなった13).したがって,TDIFが持つSer11は自身の受容体であるTDRを活性化するためには必要ではないが,類似のCLV3ペプチドの受容体であるCLV1を活性化しないように働いており,細胞間情報伝達の特異性をもたらす役割があると考えられる.ただし,CLV3のHis11をSer11に置換したペプチドはCLV3と同様にCLV1と結合することから,リガンド–受容体間の結合を阻害するのではなく,共受容体との相互作用などに影響する可能性が考えられている.この結果は植物ゲノム中に存在する遺伝的多様性がペプチドの生理活性の改変に利用できることも示唆している.
近年のゲノム研究の進展によって,陸上植物やその近縁の藻類のゲノムが解読されつつある.その結果,CLEを含む短鎖ペプチドホルモンの遺伝子が陸上植物に広く保存されていることが明らかとなった.同様に,陸上植物ではLRR型受容体様キナーゼのほぼすべてのサブクラスが共有されている14).現生の陸上植物は,維管束植物とコケ植物に大きく分けられ,さらに被子植物やシダ植物といった主要な7系統に分けることができる.系統間では根・葉・茎・維管束といった植物体の基本構造が異なるにもかかわらず,類似のペプチドホルモンを用いて植物体の成長生理を調節していると考えられる.
ゼニゴケは近年モデル生物として確立されたコケ植物の一種で,タイ類に属している.ゼニゴケの植物体は茎や葉の区別のない葉状体であり,維管束や種子,花などを作らない.しかしながら,ゼニゴケにはシロイヌナズナに存在する発生調節遺伝子群はサブファミリーレベルでおおむね保存されている.ただし,遺伝子コピー数が少ないという特徴を持つことが明らかとなっている.
筆者らはゼニゴケゲノムにおいてCLE遺伝子を探索し,2遺伝子(MpCLE1, MpCLE2)を見いだした(図3A).これは32個のCLE遺伝子を持つシロイヌナズナとは対照的であった.MpCLE1はTDIFのホモログである一方で,MpCLE2はCLV3のホモログであることから,それぞれが各サブファミリー内の単一メンバーであることが示唆された.
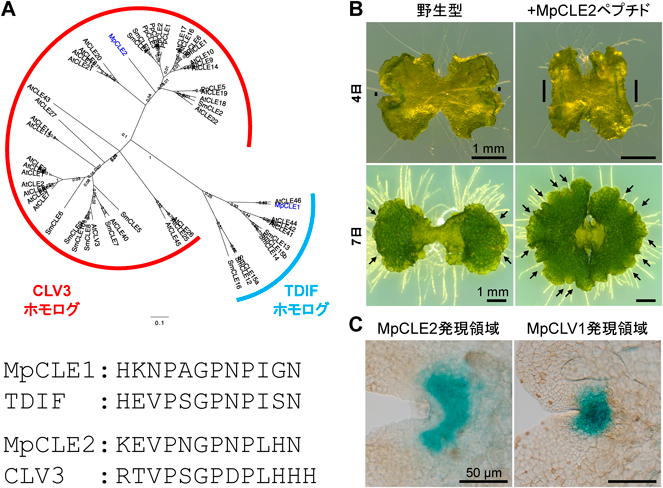
(A)ゼニゴケとシロイヌナズナのCLEペプチド群の比較.分子系統とアミノ酸配列を示す.(B)ゼニゴケMpCLE2ペプチドの作用.MpCLE2ペプチドによって,育成4日では分裂組織の幅(太線)が広がり,育成7日では多数の分裂組織(矢印)の形成が誘導される.(C)分裂組織内でのMpCLE2とMpCLV1の遺伝子発現領域の比較.文献15および16より改変.
MpCLE1はTDIFのホモログであるが,ゼニゴケは維管束を持たないため,機能が異なると予想された15).MpCLE1遺伝子の過剰発現体やノックアウト個体の解析から,MpCLE1はゼニゴケの分裂組織における細胞増殖を抑制する生理機能を持つことが示唆された.シロイヌナズナのTDIF受容体であるTDRについても,ゼニゴケには単一のホモログ(MpTDR)が存在している.この受容体の欠損変異体ではMpCLE1ペプチドに対する感受性が失われていた.ゼニゴケにシロイヌナズナのTDIFペプチドを与えた場合でもMpTDR依存的に同様の活性がみられた.したがって,TDIF–TDRのリガンド–受容体ペアはゼニゴケとシロイヌナズナの間で共通しているが,その生理機能は異なっているといえる.
MpCLE2はCLV3のホモログである16).ゼニゴケには花や果実は存在しないものの,分裂組織が存在する点ではシロイヌナズナと共通している.筆者らはMpCLE2の合成ペプチドをゼニゴケの幼植物体に与えて4日間育成したところ,分裂組織が拡大することを見いだした(図3B).この植物体をMpCLE2ペプチド非含有培地に移す実験を行ったところ,拡大した分裂組織から複数の新たな分裂組織が生じ,体軸の分岐が増えることがわかった.したがって,MpCLE2ペプチドは分裂組織の幹細胞を増やす生理機能を持つことが示唆された.受容体CLV1のホモログもゼニゴケには1遺伝子のみ存在しており,このMpCLV1遺伝子の欠損変異体ではMpCLE2に対する感受性が失われた.したがって,MpCLE2–MpCLV1のリガンド–受容体ペアもゼニゴケとシロイヌナズナの間で共通している.
分裂組織の内部において,MpCLE2を発現する領域は周縁部に位置するのに対し,MpCLV1は分裂組織中央の幹細胞群を含む領域で発現していた.したがって,分裂組織内での細胞間シグナルによって幹細胞数を調節していることが示唆される(図3C).したがって,シロイヌナズナCLV3とゼニゴケMpCLE2は,いずれも植物成長の源である分裂組織で産生され,幹細胞動態を調節する役割を担うペプチドホルモンであるといえる.
上述のように,CLV3/CLEペプチドホルモンの生理活性について被子植物シロイヌナズナとコケ植物ゼニゴケでの共通性が示唆されたものの,幹細胞維持や細胞分化の観点では両者の生理作用は逆転している.すなわち,分化を促進するCLV3が幹細胞維持の抑制因子であるのに対し,MpCLE2は幹細胞維持の促進因子である.このような幹細胞維持に対する活性の逆転は植物の生存にとっては有害と考えられ,進化的な解釈は困難であった.
一般的に,特定の遺伝子が新たな機能を獲得する際に重要な役割を担うのが遺伝子重複である.遺伝子重複によって二つの遺伝子が生じれば,元の遺伝子機能を維持しながら新しい遺伝子の機能を獲得することができるためである(図4).興味深いことに,最近,シロイヌナズナにおいても幹細胞維持を促進するCLE40ペプチドが報告された17).CLE40とCLV3は類似の構造を持つが,それぞれBAM1とCLV1という特定の受容体分子種で受容される.CLE40–BAM1とCLV3–CLV1のペアはリガンドと受容体がそれぞれホモログの関係にあるが,分裂組織内での発現部位が異なるために生体内で独立の経路として機能する(図4).
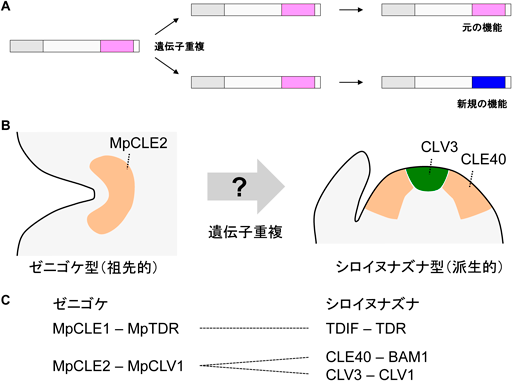
(A)遺伝子重複によるペプチドホルモンの機能進化の概念図.(B)CLEペプチドの機能進化に関するモデル.MpCLE2とCLE40は分裂組織の周縁部で発現する幹細胞維持の促進因子として共有されている.分裂組織の中央部で発現する幹細胞維持の抑制因子であるCLV3は遺伝子重複によって派生した.(C)本稿におけるCLEペプチド–受容体ペア群の類縁関係.点線は同一サブファミリー内に属すことを示す.
これらの知見を踏まえ,筆者はMpCLE2とCLE40にみられる幹細胞維持の促進活性が陸上植物に共通の普遍的な生理機能であり,幹細胞維持を阻害する古典的なCLV3ペプチドの機能は遺伝子重複によって派生的に獲得されたというモデルを提唱した18)(図4).CLV3の制御下にある幹細胞群は花や果実の源となる細胞集団であり,被子植物の祖先がこの経路を獲得したことが花や果実の進化に重要な役割を担ったのではないかと推測した.このモデルは現時点での実験的な証拠が少ないが,さまざまな陸上植物系統を解析対象とすることによって実験的な検証が可能な作業仮説である.今後の研究では,植物ペプチドホルモンの遺伝子重複が植物発生に与えた影響を解明することで,植物の成長生理の普遍的な仕組みの解明につながると期待される.
本研究に携わっていただいた多くの共同研究者の皆様に深く感謝致します.本研究の一部はJSPS科研費19K06727, 22H02676, 安倍能成記念教育基金学術研究助成金の支援を受けて行いました.
1)松林嘉克,平川有宇樹(2016)第9章ペプチドホルモン.新しい植物ホルモンの科学 第3版(浅見,柿本編),pp. 121–136, 講談社サイエンティフィク.
2) Stintzi, A. & Schaller, A. (2022) Biogenesis of post-translationally modified peptide signals for plant reproductive development. Curr. Opin. Plant Biol., 69, 102274.
3) Takahashi, F. & Shinozaki, K. (2019) Long-distance signaling in plant stress response. Curr. Opin. Plant Biol., 47, 106–111.
4) Gou, X. & Li, J. (2020) Paired receptor and coreceptor kinases perceive extracellular signals to control plant development. Plant Physiol., 182, 1667–1681.
5) Clark, S.E., Williams, R.W., & Meyerowitz, E.M. (1997) The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell, 89, 575–585.
6) Fletcher, J.C., Brand, U., Running, M.P., Simon, R., & Meyerowitz, E.M. (1999) Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science, 283, 1911–1914.
7) Cock, J.M. & McCormick, S. (2001) A large family of genes that share homology with CLAVATA3. Plant Physiol., 126, 939–942.
8) Ito, Y., Nakanomyo, I., Motose, H., Iwamoto, K., Sawa, S., Dohmae, N., & Fukuda, H. (2006) Dodeca-CLE peptides as suppressors of plant stem cell differentiation. Science, 313, 842–845.
9) Kondo, T., Sawa, S., Kinoshita, A., Mizuno, S., Kakimoto, T., Fukuda, H., & Sakagami, Y. (2006) A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science, 313, 845–848.
10) Ohyama, K., Shinohara, H., Ogawa-Ohnishi, M., & Matsubayashi, Y. (2009) A glycopeptide regulating stem cell fate in Arabidopsis thaliana. Nat. Chem. Biol., 5, 578–580.
11) Hirakawa, Y., Shinohara, H., Kondo, Y., Inoue, A., Nakanomyo, I., Ogawa, M., Sawa, S., Ohashi-Ito, K., Matsubayashi, Y., & Fukuda, H. (2008) Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proc. Natl. Acad. Sci. USA, 105, 15208–15213.
12) Hirakawa, Y. & Bowman, J.L. (2015) A role of TDIF peptide signaling in vascular cell differentiation is conserved among euphyllophytes. Front. Plant Sci., 6, 1048.
13) Hirakawa, Y., Shinohara, H., Welke, K., Irle, S., Matsubayashi, Y., Torii, K.U., & Uchida, N. (2017) Cryptic bioactivity capacitated by synthetic hybrid plant peptides. Nat. Commun., 8, 14318.
14) Furumizu, C., Krabberød, A.K., Hammerstad, M., Alling, R.M., Wildhagen, M., Sawa, S., & Aalen, R.B. (2020) The sequenced genomes of nonflowering land plants reveal the innovative evolutionary history of peptide signaling. Plant Cell, 33, 2915–2934.
15) Hirakawa, Y., Uchida, N., Yamaguchi, Y.L., Tabata, R., Ishida, S., Ishizaki, K., Nishihama, R., Kohchi, T., Sawa, S., & Bowman, J.L. (2019) Control of proliferation in the haploid meristem by CLE peptide signaling in Marchantia polymorpha. PLoS Genet., 15, e1007997.
16) Hirakawa, Y., Fujimoto, T., Ishida, S., Uchida, N., Sawa, S., Kiyosue, T., Ishizaki, K., Nishihama, R., Kohchi, T., & Bowman, J.L. (2020) Induction of Multichotomous Branching by CLAVATA Peptide in Marchantia polymorpha. Curr. Biol., 30, 3833–3840.e4.
17) Schlegel, J., Denay, G., Wink, R., Pinto, K.G., Stahl, Y., Schmid, J., Blümke, P., & Simon, R.G. (2021) Control of Arabidopsis shoot stem cell homeostasis by two antagonistic CLE peptide signalling pathways. eLife, 10, e70934.
18) Hirakawa, Y. (2022) Evolution of meristem zonation by CLE gene duplication in land plants. Nat. Plants, 8, 735–740.

学習院大学理学部生命科学科助教.博士(理学).
2006年東京大学理学部卒業.11年同大学院理学系研究科修了.11年よりMonash University博士研究員.13年より名古屋大学WPI-ITbM博士研究員.17年より現職.
研究テーマと抱負植物の分裂組織の維持調節に働くCLEペプチドホルモンを研究しています.研究を通して陸上植物の多様性や進化の解明に貢献したいと考えています.
ウェブサイトhttps://www.univ.gakushuin.ac.jp/sci/bio/laboratory/detail-kiyosue/

This page was created on 2023-03-07T14:37:38.785+09:00
This page was last modified on 2023-04-14T10:53:02.000+09:00
このサイトは(株)国際文献社によって運用されています。