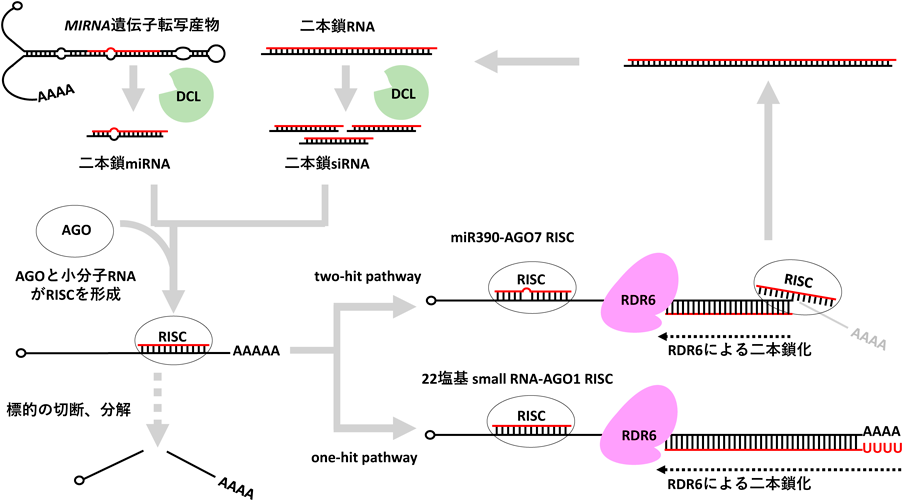
RNAサイレンシングは,小分子RNAが転写段階,あるいは転写後段階において,相補的な配列を持つ標的遺伝子の発現を負に制御する遺伝子発現制御機構である.植物の小分子RNAはその前駆体の種類によって,microRNA(miRNA),small interfering RNA(siRNA)の二つのグループに大別される.miRNAはゲノムにコードされたMIRNA遺伝子の転写産物内部のヘアピン構造部分が,DICER-LIKE1(DCL1)によるプロセシングを受けることで生成される.他方,siRNAは二本鎖RNAを前駆体とし,DCL2, DCL3, DCL4によってそれぞれ22, 24, 21塩基の長さにプロセシングされる1).生成された小分子RNAは,機能タンパク質であるARGONAUTE(AGO)タンパク質に取り込まれ,RNA-induced silencing complex(RISC)と呼ばれる複合体を形成することで,サイレンシング能を発揮する.
植物においては,標的RNAを認識したRISCにリクルートされたRNA-dependent RNA polymerase(RDR)がその標的RNAを二本鎖RNAに変換し,その二本鎖RNAをDCLがプロセシングし二次的siRNAを生成することで,効率的にRNAサイレンシングを増幅する経路が存在する.この増幅機構は分化や発生といった個体内の生命活動の維持に重要な役割を持つ他に,トランスポゾンやウイルスといった外来遺伝子への防御応答としてもきわめて重要な役割を担っている1).本稿では,最新の知見を踏まえ,この増幅経路の要であるRDRがRISCにリクルートされる分子機構について議論したいと思う.
植物の二次的siRNAには,ウイルスやトランスポゾンといった外来遺伝子に由来するものの他に,内在のものも存在し,phased siRNA(phasiRNA)はその代表的なものである.phasiRNAの前駆体はPolIIによる転写産物であり,これらのRNAはRDRによる二本鎖化の後,DCLによる21塩基あるいは24塩基ごとの等間隔な切断(phasing)を経てphasiRNAを生成する2).phasiRNAの中でもtrans-acting siRNA(tasiRNA)と呼ばれる一群はその生成機構が精力的に解析されている.
1)trans-acting siRNA
tasiRNAの生成は,ゲノム中のTAS遺伝子からTAS mRNAが転写されることに始まる.TAS mRNAが特定のmiRNAとAGOからなるRISC(22塩基のmiRNAを取り込んだAGO1またはmiR390を取り込んだAGO7)に認識されると,RDR6がリクルートされ二本鎖化が行われる.合成された二本鎖RNAは,DCL4によるプロセシングにより21塩基のtasiRNAとなり,主にAGO1とRISCを形成することで,その名のとおり自身とは異なるmRNAを標的にトランスな発現抑制を行う.tasiRNAの生成には,RISCを形成するAGOや,RDR6, DCL4の他に,二本鎖RNA結合タンパク質であるSUPPRESSOR OF GENE SILENCING 3(SGS3),機能未知のタンパク質であるSILENCING DEFECTIVE 5(SDE5)も必要であることが遺伝学的に示されている.SGS3はRISCと相互作用すること,前駆体RNAの安定化に寄与している3)ことに加え,細胞内で膜と相互作用する細胞質顆粒を形成し,RDR6やAGO7などと共局在することが報告されている4).SDE5はRISCとSGS3が相互作用してから,RDR6がリクルートされるまでのどこかの段階で何らかの機能を担っていることが示されている.
シロイヌナズナにはTAS1a/b/c, TAS2, TAS3a/b/c, TAS4の8種類のTAS遺伝子座が存在し,TAS1とTAS2のmRNAはmiR173-AGO1 RISCにより,TAS3 mRNAはmiR390-AGO7 RISCにより,TAS4 mRNAはmiR828-AGO1 RISCによりそれぞれ認識されてRDR6のリクルートが起きるが,その分子機構はRISCの結合部位の数によって「one-hit pathway」と「two-hit pathway」に分類されている.TAS3 tasiRNA生成機構のみがtwo-hit pathwayを経て生成され,それ以外のTAS1/2/4 tasiRNAおよびその他のphasiRNAはone-hit pathwayにより生成される5)(図1).
2)two-hit pathway
TAS3 mRNA上には,miR390-AGO7 RISCに認識されるmiR390と相補的な配列が2か所存在し,その両方にRISCが結合することがtwo-hit pathwayの第一段階である.第二段階では,2か所の結合サイトのうち,3′末端側の結合サイトがRISCによる切断を受ける.5′末端側の結合サイトは,中央部に保存されたミスマッチがあり,RISCの結合は起きるが切断には至らない一方,3′末端側の結合サイトの中央部は完全に相補的であるため,切断が生じる.続く第三段階では,リクルートされたRDR6がRISCにより切断された3′末端側の結合サイトから5′末端側の結合サイトに向けて相補鎖を合成する.第四段階では,合成された二本鎖がDCL4により21塩基にプロセシングされ,tasiRNAが完成する.
3)one-hit pathway
TAS1/2/4 mRNAおよびtasiRNAを除いたphasiRNAの前駆体となるmRNA上には,22塩基miRNAとAGO1により構成されるRISCに認識される配列が1か所存在する.この結合サイトはRISCによる切断を受けるが,SGS3がTAS mRNAと結合したRISCと相互作用することで,切断の後も断片どうしをつなぎ止めていることが明らかにされている3).二本鎖RNAのプロセシングを行うDCLのすべてを変異させたdcl2/3/4変異体では,TAS mRNAの3′ poly(A)配列部分にも相補鎖を持った二本鎖RNAが蓄積していることから,3′ poly(A)配列からRDR6は相補鎖合成を開始できると考えられる6)が,リコンビナントRDR6を用いた生化学的解析では,RDR6はpoly(A)を持たないRNAを鋳型としてより好むことが報告されている7).
二次的siRNA生成経路は,phasiRNAのみならず,ウイルスやトランスポゾンのような外来遺伝子に対する防御機構としてもきわめて重要であり,おそらくはtasiRNA生成経路と多くの部分を共有していると考えられる.しかしその重要性に比して,詳細な分子機構はあまりにも未解明な点が多い.では,何がその解明を阻んでいたのであろうか.障壁の一つは,生化学的な実験系の欠如であったといえる.RISC, SGS3, SDE5, RDR6,前駆体RNAといった複数の因子が関わる複雑な反応について,個々の因子がどのようなタイミングでどのように関わっているのかを明らかにするためには,tasiRNA生成経路を試験管内で再構成できる系を確立した上で,各因子の変異体を用いた解析や,それらを添加するタイミングの検討などを行う必要がある.以下,試験管内tasiRNA生成系を用いた解析による,複数の新規知見を概説する.
3. 試験管内TAS3 tasiRNA生成経路の再構成とその評価
Sakuraiらは,タバコ培養細胞Bright Yellow-2(BY-2)細胞由来の抽出液を用いて,試験管内でのTAS3 tasiRNA生成経路の再構成を試みた8).本抽出液は,高い翻訳活性を持ち,AGO1 mRNAとmiRNAの二本鎖を加えることで機能的なRISC形成の再現ができること9),DCL活性を持つことが報告されている.この抽出液にシロイヌナズナ由来のtasiRNA生成関連因子であるAtAGO7 mRNA, AtSDE5 mRNA, AtSGS3 mRNA, AtRDR6 mRNA, TAS3 mRNA, miR390/miR390*を加えたところ,TAS3 tasiRNAが蓄積した.これらの因子のうち,AtSGS3 mRNAとAtRDR6 mRNAは加えなくてもtasiRNAが十分量蓄積したことから,抽出液中には一定量の内在のSGS3とRDR6が存在し,これらはAtSGS3, AtRDR6のTAS3 tasiRNA生成における機能を代替するのに十分であると考えられた.一方,AtAGO7 mRNA, AtSDE5 mRNA, TAS3 mRNA, miR390/miR390*を加えなかった場合ではtasiRNAは蓄積しなかったため,以降の実験はこれら4因子を抽出液に添加して行った.
植物個体内では,21塩基ごとに切断を受けて生成されたtasiRNAは,それぞれが同程度に蓄積するのではなく前駆体mRNAの特定の領域から発現したもののみが高蓄積しており,この領域をhotspotと呼ぶ5).試験管内で生成したtasiRNAをRNAシークエンスしマッピングしたところ,植物個体でみられるhotspotと共通した領域にhotspotが確認されたことから,この試験管内再構成系は植物個体におけるTAS3 tasiRNA生成系を基本的に再現できていると考えられた.
4. SGS3とSDE5は協調してRDR6のリクルートに関わる
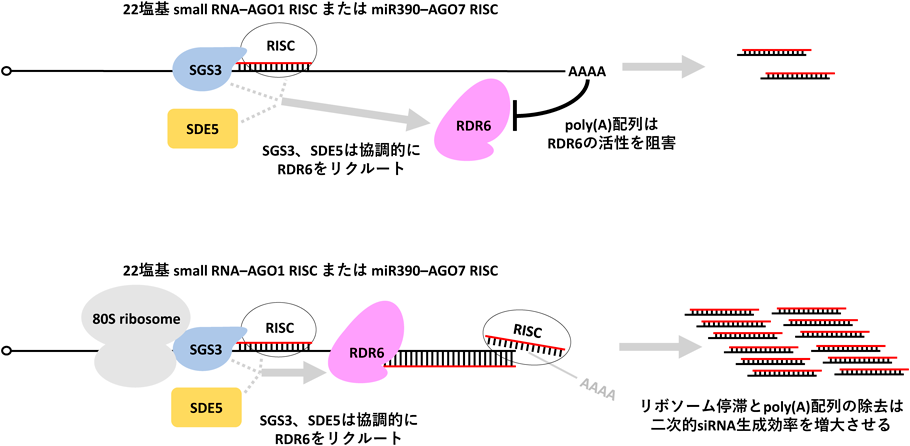
SGS3とSDE5の役割を解析するため,再構成系からこれらの因子を抜いた場合について試験を行ったところ,TAS3 mRNAのRISCによる切断は問題なく起きていた一方で,RDR6による二本鎖化が起きていないことがわかった.Sakuraiらはこの結果から,SGS3とSDE5はRISCと複合体を形成してRDR6のリクルートに関わっていると仮説を立て,共免疫沈降によりこれを検証した.結果,SGS3またはSDE5が十分量存在しない条件下では,RISCとRDR6との相互作用が起こらなかったことから,これら2因子は協調してRISCを介してTAS3 mRNAにRDR6をリクルートする役割があることが明らかとなった.
5. TAS3 two-hit pathwayにおいて,2か所のRISC結合サイトは独立な役割を担う
TAS3 mRNA上に2か所存在するRISC結合サイトの機能を解析するため,5′末端側の結合サイト,3′末端側の結合サイトおよびその両方をRISCの結合が起こらないような配列に置換した変異体TAS3 mRNAについて,RISCとRDR6との相互作用を共免疫沈降で確認した.結果,RISCとRDR6との相互作用は5′末端側の結合サイトにのみ依存していた.興味深いことに,3′末端側の結合サイトのみを変異させたTAS3 mRNAでは,RISCとRDR6の相互作用は起きているにもかかわらずRDR6による二本鎖化が起きていなかった.
リコンビナントRDR6が3′ poly(A)配列を持たない一本鎖RNAを相補鎖合成の鋳型としてより好むことから,3′末端側の結合サイトの意義は,TAS3 mRNAの切断を引き起こしpoly(A)配列を除去することにあると仮説を立てたSakuraiらは,3′末端側結合サイトの変異体TAS3 mRNAの3′ poly(A)配列を非poly(A)配列に置換することで,二本鎖化が起こるのではないかと考えた.そこで,実際にpoly(A)配列を持つ場合と非poly(A)配列に置換した場合について,RISCとRDR6との相互作用とTAS3 mRNA二本鎖化を解析したところ,相互作用は同程度であった一方,非poly(A)配列置換を行った場合に特異的に二本鎖化が起こった.以上より,3′末端側の結合サイトは3′ poly(A)配列を除去し,RDR6の活性を高める役割を担っていることが明らかとなった.
6. one-hit pathwayにおいては22塩基のsmall RNAとAGO1のRISCが二次的siRNA産生を引き起こす
one-hit pathwayの分子機構についてはシロイヌナズナの培養細胞由来の抽出液を用いた生化学的解析を行ったYoshikawaらの報告が詳しい10).Yoshikawaらは,抽出液にtasiRNA生成能があることを確認した後,人工的に設計したレポーター系を用い,21塩基のsmall RNAとAGO1が形成するRISCと22塩基のsmall RNAとAGO1が形成するRISCのどちらが二次的siRNA合成を引き起こすのか解析した.結果,22塩基のsmall RNAとAGO1で形成されたRISCがより強く二次的siRNA合成を引き起こすことがわかった.
Yoshikawaらのone-hit pathwayをモデルとする実験系においても,RDR6はSGS3とSDE5の両者の存在下でのみ標的RNAにリクルートされた.また,3′ poly(A)配列を持つ標的RNAはRDR6による二本鎖化の効率が低かった.したがって,SGS3とSDE5が協調してリクルートしたRDR6が,非3′ poly(A)配列を好んで二本鎖化し二次的siRNA生成を引き起こすというモデルは,two-hit pathwayとone-hit pathwayに共通するものであると考えられた.one-hit pathwayにおける二本鎖化はRISC結合サイトの3′末端側で起きているため,実際には標的RNAは3′ poly(A)配列を持つ場合が想定される.確かに3′ poly(A)配列からRDR6が相補鎖合成を開始することは可能ではあるが,植物細胞内では,先に生成されていたtasiRNAによるTAS mRNAへのセルフアタックにより3′ poly(A)配列を含む3′末端配列が除去される,two-hit pathwayのような過程によりRDR6の働きが促進されている可能性がある.
7. SGS3はRISCと協調してリボソーム停滞を引き起こし,二次的siRNA産生を促進する
植物においては,多くのmiRNAは21塩基であり,AGO1とRISCを形成して標的のサイレンシングを行う.一方,二次的siRNA産生を引き起こすRISCは22塩基のmiRNAとAGO1,またはmiR390とAGO7により構成されたものに限られている.これらの限定的なRISCのみが二次的siRNA産生を誘導する能力を持つのはなぜであろうか.Wuらは,環境ストレスに応じて蓄積する22塩基のsiRNAは,標的の切断を伴わずに翻訳抑制すること,二次的siRNAの蓄積を誘導することを見いだした11).siRNAではなくmiRNAにおいても,人工的に設計したRNAを用いた解析により,miR173結合部位をORFの直後に配置した場合に二次的siRNA産生が促進されることを見いだした報告12)や,TAS2 mRNAのmiR173結合部位にオーバーラップした短いORFが翻訳されることがtasiRNA生成に重要であることを示した報告13)がなされている.
これらの22塩基のsmall RNAが植物体において翻訳抑制を引き起こし二次的siRNA合成の引き金となるというこれまでの知見は,Yoshikawaらによる22塩基のsmall RNAとAGO1によるRISCがより多くの二次的siRNA産生を引き起こすという報告ともよく調和する.それでは,この翻訳抑制はどのように二次的siRNA合成とつながっているのだろうか.そして,このつながりはone-hit pathwayだけではなく,two-hit pathwayにも共通しているのだろうか.
TAS1 mRNA, TAS2 mRNAのRISC結合サイトは短いORFと重複している.この短いORFはtasiRNA生成に重要であることが示されており,TAS3 mRNAのRISC結合サイトの直前にもORFが存在している.シロイヌナズナの植物体を用いたリボソームプロファイリングの結果,TAS mRNA上のRISC結合サイトの直上に大量のリードが蓄積していたことから,ここでリボソームの停滞が起きていると考えられた.sgs3変異体シロイヌナズナについて同様にリボソーム停滞を解析すると,TAS mRNA上のリボソーム停滞は起きていなかったため,TAS mRNA上のリボソーム停滞にはSGS3が何らかの必須な役割を担っていると考えられた.BY-2細胞由来抽出液を用いた無細胞系での解析から,SGS3はmiR390または22塩基small RNAの3′末端を認識しTAS mRNA, RISCに直接結合していることが明らかとなった14).
TAS3mRNAのRISC結合サイトから段階的に直上のORFを遠ざけ,リボソームとRISC-SGS3複合体との衝突が起こらないようにした変異体TAS3 mRNAをNicotiana benthamianaに発現させtasiRNAの蓄積を解析すると,リボソーム停滞の起こらない変異体からのtasiRNAの蓄積は大きく減少していた.以上より,RISCがTAS mRNAに結合すると,small RNAの3′末端を介してSGS3がRISC, TAS mRNAと複合体を形成し,リボソーム停滞を引き起こすことで二次的siRNA産生を促進するというモデルが提示された14)(図2).
本稿では,植物の二次的siRNA生成機構について概説した.一連の研究により,標的RNAを認識したRISCがSGS3, SDE5と協調しRDRをリクルートすることが明らかとなったが,これらの因子がどのようにリクルートに関与しているのかの詳細は依然として明らかになっていない.同様に,poly(A)配列がRDRの活性を抑制する機構や,リボソーム停滞が二次的siRNA生成を促す機構も詳細は不明である.加えて近年,SGS3のN末端側に存在するプリオン様ドメインが液-液相分離を引き起こし,細胞内でsiRNA bodyと呼ばれる液滴を形成することが報告されている.siRNA bodyにはRDR6, AGO7, SDE5なども含まれており二次的siRNA生成における重要な役割を担っていると示唆されている15).SGS3の機能解析を含む多角的な解析により,未解明な点を含む二次的siRNA生成機構のさらに詳細な理解が進むことが期待される.
引用文献References
1) Bologna, N.G. & Voinnet, O. (2014) The diversity, biogenesis, and activities of endogenous silencing small RNAs in Arabidopsis. Annu. Rev. Plant Biol., 65, 473–503.
2) Liu, Y., Teng, C., Xia, R., & Meyers, B.C. (2020) PhasiRNAs in plants: Their biogenesis, genic sources, and roles in stress responses, development, and reproduction. Plant Cell, 32, 3059–3080.
3) Yoshikawa, M., Iki, T., Tsutsui, Y., Miyashita, K., Poethig, R.S., Habu, Y., & Ishikawa, M. (2013) 3′ fragment of miR173-programmed RISC-cleaved RNA is protected from degradation in a complex with RISC and SGS3. Proc. Natl. Acad. Sci. USA, 110, 4117–4122.
4) Jouannet, V., Moreno, A.B., Elmayan, T., Vaucheret, H., Crespi, M.D., & Maizel, A. (2012) Cytoplasmic Arabidopsis AGO7 accumulates in membrane-associated siRNA bodies and is required for ta-siRNA biogenesis: AGO7 associates with membranes. EMBO J., 31, 1704–1713.
5) Axtell, M.J., Jan, C., Rajagopalan, R., & Bartel, D.P. (2006) A two-hit trigger for siRNA biogenesis in plants. Cell, 127, 565–577.
6) Rajeswaran, R., Aregger, M., Zvereva, A.S., Borah, B.K., Gubaeva, E.G., & Pooggin, M.M. (2012) Sequencing of RDR6-dependent double-stranded RNAs reveals novel features of plant siRNA biogenesis. Nucleic Acids Res., 40, 6241–6254.
7) Baeg, K., Iwakawa, H., & Tomari, Y. (2017) The poly(A) tail blocks RDR6 from converting self mRNAs into substrates for gene silencing. Nat. Plants, 3, 17036.
8) Sakurai, Y., Baeg, K., Lam, A.Y.W., Shoji, K., Tomari, Y., & Iwakawa, H. (2021) Cell-free reconstitution reveals the molecular mechanisms for the initiation of secondary siRNA biogenesis in plants. Proc. Natl. Acad. Sci. USA, 118, e2102889118.
9) Iki, T., Yoshikawa, M., Nishikiori, M., Jaudal, M.C., Matsumoto-Yokoyama, E., Mitsuhara, I., Meshi, T., & Ishikawa, M. (2010) In vitro assembly of plant RNA-induced silencing complexes facilitated by molecular chaperone HSP90. Mol. Cell, 39, 282–291.
10) Yoshikawa, M., Han, Y.-W., Fujii, H., Aizawa, S., Nishino, T., & Ishikawa, M. (2021) Cooperative recruitment of RDR6 by SGS3 and SDE5 during small interfering RNA amplification in Arabidopsis. Proc. Natl. Acad. Sci. USA, 118, e2102885118.
11) Wu, H., Li, B., Iwakawa, H.O., Pan, Y., Tang, X., Ling-Hu, Q., Liu, Y., Sheng, S., Feng, L., Zhang, H., et al. (2020) Plant 22-nt siRNAs mediate translational repression and stress adaptation. Nature, 581, 89–93.
12) Zhang, C., Ng, D.W.-K., Lu, J., & Chen, Z.J. (2012) Roles of target site location and sequence complementarity in trans-acting siRNA formation in Arabidopsis. Plant J., 69, 217–226.
13) Yoshikawa, M., Iki, T., Numa, H., Miyashita, K., Meshi, T., & Ishikawa, M. (2016) A short open reading frame encompassing the MicroRNA173 target site plays a role in trans-acting small interfering RNA biogenesis. Plant Physiol., 171, 359–368.
14) Iwakawa, H.O., Lam, A.Y.W., Mine, A., Fujita, T., Kiyokawa, K., Yoshikawa, M., Takeda, A., Iwasaki, S., & Tomari, Y. (2021) Ribosome stalling caused by the Argonaute-microRNA-SGS3 complex regulates the production of secondary siRNAs in plants. Cell Rep., 35, 109300.
15) Tan, H., Luo, W., Yan, W., Liu, J., Aizezi, Y., Cui, R., Tian, R., Ma, J., & Guo, H. (2023) Phase separation of SGS3 drives siRNA body formation and promotes endogenous gene silencing. Cell Rep., 42, 111985.
著者紹介Author Profile
 藤本 祐司(ふじもと ゆうじ)
藤本 祐司(ふじもと ゆうじ)立教大学理学部岩川研究室所属日本学術振興会特別研究員.博士(農学).
略歴2017年東京大学農学部卒業.22年3月東京大学大学院農学生命科学研究科博士課程修了.同年4月より現職.
研究テーマと抱負植物におけるmRNAの制御機構を包括的に把握することを最終的な目標としながら,現在は主に二次的siRNA生成機構とmRNA代謝機構の関わりあいについて関心を持って研究している.
ウェブサイトhttps://sites.google.com/rikkyo.ac.jp/iwakawalab/home
趣味暗渠になった古い川の跡を探す.
 岩川 弘宙(いわかわ ひろおき)
岩川 弘宙(いわかわ ひろおき)立教大学理学部生命理学科准教授.博士(農学).
略歴2005年京都大学農学部卒業.10年同大学院農学研究科博士課程修了.以後,日本学術振興会特別研究員,東京大学分子細胞生物学研究所助教,JSTさきがけ研究者,東京大学定量生命科学研究所講師等を経て,22年より現職.
研究テーマと抱負小分子RNAが関わるRNAサイレンシング機構の理解.
ウェブサイトhttps://sites.google.com/rikkyo.ac.jp/iwakawalab/home
趣味築地で仕入れた新鮮な食材を使っての料理.