高速運動する植物ミオシンFast-moving plant myosins
千葉大学大学院理学研究院生物学研究部門Department of Biology, Graduate School of Science, Chiba University ◇ 〒263–8522 千葉市稲毛区弥生町1–33 ◇ 1–33 Yayoi-cho, Inage, Chiba 263–8522, Japan
発行日:2023年6月25日Published: June 25, 2023
© 2023 公益社団法人日本生化学会© 2023 The Japanese Biochemical Society
植物ミオシンは動物ミオシンと比べて桁違いの高い運動速度を持つ.我々はこれまで,タンパク質的アプローチを中心に植物ミオシンの解析を行ってきた.これまでの研究により,植物ミオシンの高速運動を引き起こす構造的基盤1, 2)酵素的基盤3, 4),高速運動の生理的意義5),植物ミオシンの運動速度多様性6–8),植物ミオシンの調節機構9)など高速運動する植物ミオシンの特性を明らかにしてきた.本稿では,我々の研究成果を中心に高速運動する植物ミオシンについて概説する.
ミオシンはATPの加水分解エネルギーを利用して,アクチン繊維上を運動する代表的なモータータンパク質であり,ほぼすべての真核生物細胞に存在する.ミオシンは動物において,走る・飛ぶ・跳ねるなどの個体レベルの運動の他,心臓拍動・血管収縮などの器官レベルの運動,細胞運動・細胞質分裂などの細胞レベルの運動,細胞内物質輸送などの細胞内レベルの運動などさまざまなレベルの運動に関わっている.ミオシンは1942年のSzent-Györgyiによる筋組織の収縮研究で発見された.そして,発見後の約50年間は,ミオシン研究のほとんどは筋肉ミオシンを用いて行われてきた.1990年代から2000年にかけてのゲノム解析の結果,ヒトには39遺伝子のミオシンが存在することがわかった.このうち,14遺伝子は骨格筋,心筋,平滑筋といった筋肉ミオシンが属する「ミオシン2」であったが,25遺伝子は筋肉ミオシンとは異なるクラスのミオシンであることがわかった.それらは発見順にミオシン1, 3, 5, 6, 7, 9, 10, 15, 16, 18と名づけられた.ちなみに,ミオシン1は筋肉ミオシンであるミオシン2の30年後に発見されたが,筋肉ミオシンがcoiled-coilにより二量体を形成しているのに対し,ミオシン1はcoiled-coilがなく単量体であったので,先に発見された二量体である筋肉ミオシンがミオシン2と名づけられ,後に発見された単量体のミオシンがミオシン1と名づけられた.その後,ミオシン1以外にも多くの非筋ミオシンが発見されたが,それらは,モータードメインのアミノ酸配列の相同性によるクラス分けがなされ,前述したように,発見された順に数字がつけられていった.近年,ヒト以外のさまざまな生物でゲノムプロジェクトが行われ,ミオシンは79以上のクラスからなるスーパーファミリーを構成しており,さらにそれぞれのクラスにはさまざまなサブクラスが存在していることが明らかになっている10).ミオシンはクラス,サブクラスが異なると運動速度,運動方向,力の大きさなどミオシン特性が大きく異なる11).
動物は食べ物を求めて活発に動き回る.その運動の駆動力は筋肉ミオシンである.一方,植物は光合成によりエネルギーを作ることができるので,動く必要がなく,発生した場所で一生過ごす.そのため,一般に動物は「動」,植物は「静」のイメージがある.しかし,植物の細胞内を顕微鏡で観察すると,原形質流動と呼ばれている動物細胞ではみられない激しい運動が起こっていることがわかる.原形質流動は植物特有のミオシンであるミオシン11により駆動されている.ミオシン11は球状尾部ドメイン(globular tail domain:GTD)で小胞体に結合しながら,アクチン繊維をプラス端方向に運動する(図1A, B).植物細胞の細胞膜直下ではアクチン繊維が極性をそろえて配向しているので,植物細胞内では一方向性の流れである原形質流動が発生している(図1C).
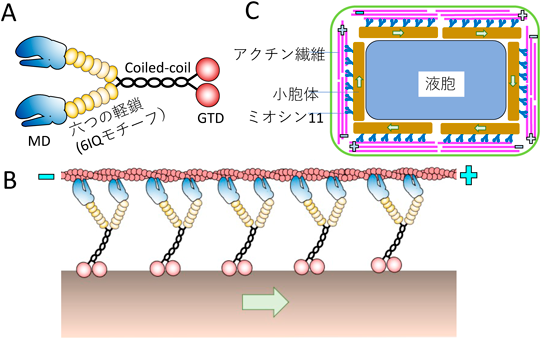
(A)ミオシン11の模式図.N末端にATPおよびアクチン繊維と結合するモータードメイン(MD)が存在する.MDのC末端側にはカルモジュリンもしくはカルモジュリン様タンパク質が結合するIQモチーフが六つあり,六つの軽鎖(カルモジュリンもしくはカルモジュリン様タンパク質)が結合する.六つの軽鎖が結合した6IQはミオシン運動における「レバーアーム」として機能する.IQモチーフのC末端側にはcoiled-coil領域が存在し,これによりミオシン11は二量体を形成する.C末端側には小胞体をはじめさまざまなオルガネラと結合す球状尾部ドメイン(GTD)が存在する.(B)ミオシン11は球状尾部ドメインで小胞体に結合し,頭部のモータードメインでアクチン繊維のプラス端方向に運動する.(C)植物細胞内の細胞膜直下ではアクチン繊維が極性をそろえて配向しており,小胞体に結合したミオシン11がアクチン繊維のプラス方向に運動することによる流体力学的効果により,植物細胞内では原形質流動が起きている.
原形質流動は非常にダイナミックな運動であるので創成期の簡易的な顕微鏡でも容易に観察でき,文献としての原形質流動の最初の報告は1774年のイタリアのCortiによる淡水産藻類のシャジクモの原形質流動観察まで遡る.ちなみに,植物の中で最も速い原形質流動を行っている生物は最初に原形質流動が報告されたシャジクモであり,その速度は25°Cで70 µm/sに及ぶ.シャジクモの原形質流動はその高速運動に加え,非常にきれいな一方向性の様相を呈するので,原形質流動の研究材料として古くから使用されてきた.1950年代に神谷・黒田は,シャジクモ細胞の原形質流動速度は細胞膜近傍が最も速く,細胞膜から離れるほど遅くなることを見いだし,流体力学的考察から原形質流動の駆動装置が細胞膜直下に存在することを提唱した12).1974年にシャジクモの細胞膜直下にアクチン繊維が極性をそろえて配向していることがわかり13),原形質流動の駆動はミオシンによると考えられるようになった.このことはまた,シャジクモには70 µm/sの生物界最速のミオシンが存在することを意味する.多くの研究者がシャジクモからの最速ミオシンの単離を試みたが,タンパク質分解酵素に富む液胞が細胞質の大半を占めるシャジクモから生化学的な方法で純度よいミオシンを精製するのは難しく,さらに,シャジクモのゲノム情報も不足していたこともあり,1974年の最速ミオシンの存在予測から,クローニング・同定まで長い時間を要した.2018年に西山・坂山らにより明らかにされたシャジクモChara brauniiのゲノム情報14)をもとにして,我々は西山・坂山と共同でシャジクモC. brauniiから4種のミオシン11遺伝子をクローニングし,Cb11-1, 2, 3, 4と名づけた.次いで,これらの遺伝子を昆虫培養細胞の系で発現させ,精製し,in vitro運動アッセイによりそれらの速度を測定したところ,Cb11-1とCb11-2が長年にわたりその存在が予見されていた速度70 µm/sの超高速ミオシンであり,Cb11-1が73 µm/sの生物界最速のミオシンであることがわかった1).その存在が予想された1974年から論文報告の2022年まで,最速ミオシンを同定するのに実に半世紀もの時間がかかったのである.Cb11-1の速度の73 µm/sは動物のさまざまなミオシンより10~1000倍速く,動物の最速ミオシンである骨格筋ミオシンと比べても10倍速い.Cb11-1は文字どおり桁違いの速度を持つ生物界最速のミオシンである(図2).なお,樫山らにより先にシャジクモChara corallinaから遺伝子クローニングされた運動速度22 µm/sのCc1115)は,今回,我々がChara brauniiからクローニングしたCb11-3のオルソログであった.我々はまた,モデル植物として使われている双子葉植物のシロイヌナズナの13種のミオシン11についても,昆虫細胞で発現,精製し,その速度を測定した.それらの速度は多様であったが,多くは7 µm/s以上であり,植物のミオシン11は動物のどのミオシンと比べても高速であることがわかった(図2).
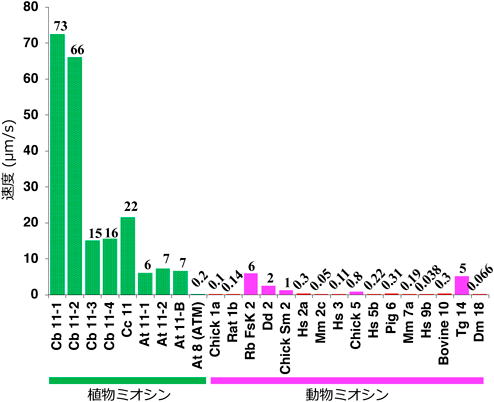
緑:植物ミオシン,マゼンタ:動物ミオシン.文献1より一部改変転載.
ミオシン11の高速運動の生理学的理由は何であろうか? その鍵は植物の細胞の大きさと関係があると考えられている.動物細胞は直径10 µm程度であるが,植物細胞は分裂を伴わない細胞成長が起こるので,成熟した細胞では直径100 µmを超え,1 mm以上の細胞もある.ブラウン運動による拡散時間は距離の二乗に比例する.そのため,直径10 µmの動物細胞においては細胞外から取り入れた糖などの低分子がブラウン運動で細胞内に拡散するのに要する時間はわずか0.05秒程度であるが,直径1 mmの植物細胞では約10分かかる.細胞サイズが大きい植物細胞ではブラウン運動による拡散だけでは不十分であるので,原形質流動により拡散を促進していると考えられる.この仮説を検証するために,我々は富永らと共同で,シロイヌナズナの原形質流動を駆動している速度7 µm/sのAt11-2のモータードメイン(MD)を速度22 µm/sのCc11のMDに置換した高速型At11-2キメラミオシンをシロイヌナズナAt11-2欠失株で発現させた.高速型At11-2キメラミオシンを導入したシロイヌナズナ株では,細胞サイズが大きくなり,それとともに植物体サイズも大きくなり,上記の仮説を支持する結果となった5).動けない植物は液胞に栄養物を貯める必要がある.このとき,液胞の増大に伴い細胞サイズも大きくなる.大きな細胞サイズでも栄養物や酸素を細胞内に行きわたらせるために,高速の原形質流動が必要となり,ミオシン11が高速に進化したのだろう.シャジクモ細胞は直径が1 mm,長さが5~10 cmであり,他の植物の細胞と比べて桁違いに大きいので,シャジクモはこのような桁違いの速さのミオシン11を持つように進化したと考えられる.もしくは,桁違いの速さのミオシン11を持つことにより,シャジクモは巨大な細胞サイズとなったのかもしれない.
最後にミオシン11の高速運動を引き起こす分子・構造基盤について我々の研究結果を概説する.ミオシンの運動速度は(ステップサイズ)/(アクチンとの強い結合時間)で近似される.ミオシンの速度は生物種,ミオシンクラスにより大きく異なり,その差は最大1000倍以上あるが,ステップサイズの違いは大きくても数倍程度であり,ミオシンの速度を主に規定するのはアクチンとの強い結合時間である.アクチンとの強い結合時間は主としてアクチン結合時のミオシンからのADP解離速度(k-ADP of acto-M)で規定される.我々はシャジクモCc11の(k-ADP of acto-M)をストップドフロー装置で測定したところ,これまで測定されたどのミオシンよりも高い値であることを見いだした(図3A, k-ADP of acto-M).興味深いことに,アクチン非結合時におけるミオシンからのADP解離速度(k-ADP of M)については,Cc11の値は他のさまざまな速度のミオシンと変わらなかった(図3A, k-ADP of M).つまり,速いミオシンはアクチン結合によってADP解離反応が促進されるが,遅いミオシンはあまり促進されない(図3A, Acceleration of ADP dissociation by actin)3).これらの結果から速いミオシンと遅いミオシンの違いはアクチンとの結合様式の違いから生じる可能性が考えられる.
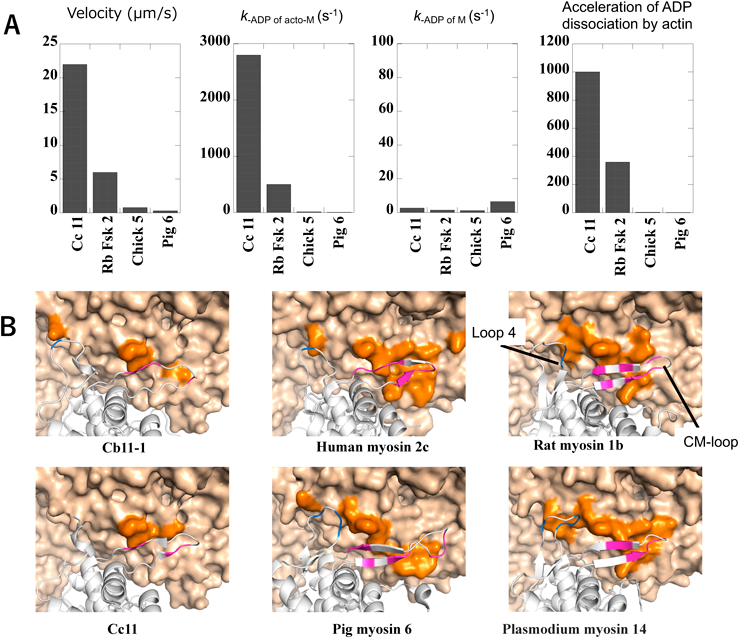
(A) Velocity:四つのクラスのミオシンの運動速度,k-ADP from acto-M:アクチンに結合した状態のミオシンからのADP解離速度,k-ADP from M:アクチン非結合時のミオシンからのADP解離速度,Acceleration of ADP dissociation by actin:アクチン結合によるミオシンからのADP解離速度の加速倍率.Cc 11:Chara corallina myosin 11, Rb Fsk 2:rabbit fast skeletal muscle myosin 2, Chick 5:chicken myosin 5, Pig 6:pig myosin 6. データは文献3より.(B)さまざまなクラスのミオシンのloop4およびCM-loopにおけるアクチンとの結合様式.濃いオレンジ色で示したloop4およびCM-loopが結合しているアクチン領域はミオシンのクラス,サブクラスにより異なっている.Cb11-1:Chara braunii myosin 11-1, Cc11:Chara corallina myosin 11. 文献1より一部改変転載.
この仮説については,以下のようにシャジクモミオシンを使った変異実験からも支持された.ミオシンのアクチン結合領域はloop2, loop3, loop 4, CM loop, helix-turn-helixの5か所から構成されている.多くのミオシンはloop2に正電荷アミノ酸が多くあり,この部分がアクチンのN末端の負電荷に富む領域と結合する.一方,シャジクモミオシンはloop2に正電荷アミノ酸がほとんどなく,アクチンとの結合に関与していなかった.その代わり,シャジクモミオシンはloop3の正電荷が他のミオシンと比べて多く,loop2の代わりとなってアクチンと結合していた.ミオシン変異実験から,loop2でアクチンに結合せず,loop3で結合するというアクチンとの結合様式の違いが,シャジクモミオシンの高速化の一因となっていることがわかった2).
我々は村田・鈴木と共同で最も高速な運動速度を持つミオシンクラスであるミオシン11の最初のX線結晶構造解析による高分解能構造として,シロイヌナズナミオシンAt11-2の高分解能(2.8 Å)構造を取得することに成功した1).At11-2のホモロジーモデルによりCb11-1とCc11の構造を予測した.そして,さまざまなミオシンの構造と比較した結果,ミオシンとアクチンとの結合は,ミオシン側の結合loop配列がミオシン間で大きく異なるだけでなく,それらのloop配列と結合するアクチン領域もミオシン間で大きく異なることがわかった(図3B).シャジクモミオシンの中でもCb11-1とCc11は速度が約3倍違うが,この二つのミオシン間ではloop4およびCM loopが結合するアクチン領域も異なっていることがホモロジーモデルから予想された(図3B).さらに,ミオシン変異実験により,loop4およびCM loopの結合様式の違いがCb11-1とCc11との間の速度の違いの一因となっていることがわかった1).
高速ミオシンの構造的知見および変異実験から,アクチンとの結合様式の多様性がミオシンの速度の多様性を生み出していることがわかった.我々は以前,植物内のミオシンを高速化すると,原形質流動が高速化し,植物の大型化につながることを示した5).ミオシンのアクチン結合部位の改変によるミオシン高速化はバイオマス・資源植物などの増産の他,高速運動するナノマシンの開発への応用も期待できる.
1) Haraguchi, T., Tamanaha, M., Suzuki, K., Yoshimura, K., Imi, T., Tominaga, M., Sakayama, H., Nishiyama, T., Murata, T., & Ito, K. (2022) Discovery of ultrafast myosin, its amino acid sequence, and structural features. Proc. Natl. Acad. Sci. USA, 119, e2120962119.
2) Ito, K., Yamaguchi, Y., Yanase, K., Ichikawa, Y., & Yamamoto, K. (2009) Unique charge distribution in surface loops confers high velocity on the fast motor protein Chara myosin. Proc. Natl. Acad. Sci. USA, 106, 21585–21590.
3) Ito, K., Ikebe, M., Kashiyama, T., Mogami, T., Kon, T., & Yamamoto, K. (2007) Kinetic mechanism of the fastest motor protein, Chara myosin. J. Biol. Chem., 282, 19534–19545.
4) Ito, K., Kashiyama, T., Shimada, K., Yamaguchi, A., Awata, J., Hachikubo, Y., Manstein, D.J., & Yamamoto, K. (2003) Recombinant motor domain constructs of Chara corallina myosin display fast motility and high ATPase activity. Biochem. Biophys. Res. Commun., 312, 958–964.
5) Tominaga, M., Kimura, A., Yokota, E., Haraguchi, T., Shimmen, T., Yamamoto, K., Nakano, A., & Ito, K. (2013) Cytoplasmic streaming velocity as a plant size determinant. Dev. Cell, 27, 345–352.
6) Haraguchi, T., Ito, K., Duan, Z., Rula, S., Takahashi, K., Shibuya, Y., Hagino, N., Miyatake, Y., Nakano, A., & Tominaga, M. (2018) Functional diversity of class XI myosins in Arabidopsis thaliana. Plant Cell Physiol., 59, 2268–2277.
7) Haraguchi, T., Tominaga, M., Nakano, A., Yamamoto, K., & Ito, K. (2016) Myosin XI-I is mechanically and enzymatically unique among Class-XI myosins in arabidopsis. Plant Cell Physiol., 57, 1732–1743.
8) Haraguchi, T., Tominaga, M., Matsumoto, R., Sato, K., Nakano, A., Yamamoto, K., & Ito, K. (2014) Molecular characterization and subcellular localization of arabidopsis Class VIII myosin, ATM1. J. Biol. Chem., 289, 12343–12355.
9) Haraguchi, T., Ito, K., Morikawa, T., Yoshimura, K., Shoji, N., Kimura, A., Iwaki, M., & Tominaga, M. (2022) Autoregulation and dual stepping mode of MYA2, an Arabidopsis myosin XI responsible for cytoplasmic streaming. Sci. Rep., 12, 3150.
10) Kollmar, M. & Muhlhausen, S. (2017) Myosin repertoire expansion coincides with eukaryotic diversification in the Mesoproterozoic era. BMC Evol. Biol., 17, 211.
11) Hartman, M.A. & Spudich, J.A. (2012) The myosin superfamily at a glance. J. Cell Sci., 125, 1627–1632.
12) Kamiya, N. & Kuroda, K. (1956) Velocity distribution of the protoplasmic streaming in Nitella cells. Shokubutsugaku Zasshi, 69, 544–554.
13) Williamson, R.E. (1974) Actin in the alga, Chara corallina. Nature, 248, 801–802.
14) Nishiyama, T., Sakayama, H., de Vries, J., Buschmann, H., Saint-Marcoux, D., Ullrich, K.K., Haas, F.B., Vanderstraeten, L., Becker, D., Lang, D., et al. (2018) The chara genome: Secondary complexity and implications for plant terrestrialization. Cell, 174, 448–464.e24.
15) Kashiyama, T., Kimura, N., Mimura, T., & Yamamoto, K. (2000) Cloning and characterization of a myosin from characean alga, the fastest motor protein in the world. J. Biochem., 127, 1065–1070.

千葉大学大学院理学研究院生物学研究部門 教授.博士(理学).
1994年名古屋大学大学院理学研究科満了,博士(理学),産総研研究員,千葉大学助手,助教,講師,准教授をへて,2015年より現職.
研究テーマと抱負千葉大学に赴任してから植物ミオシンの研究始めました.何故,植物ミオシンか?というと,それは植物ミオシンの超高速運動は見ているだけで面白い!からです.
ウェブサイトhttp://www.bio.s.chiba-u.ac.jp/ito.html
趣味野球観戦.

千葉大学大学院理学研究院生物学研究部門 助教.博士(理学).
2014年千葉大学大学院博士後期課程融合科学研究科ナノサイエンス・ナノバイオロジー修了.同年九州工業大学情報工学部研究職員.15年より千葉大学大学院理学研究院特任研究員を経て22年より現職.
研究テーマと抱負植物特異的なミオシンの機能解析.どんな時でも楽しく研究に取り組んでいきたい.
趣味ひなたぼっこ.

This page was created on 2023-05-22T08:58:04.954+09:00
This page was last modified on 2023-06-15T15:58:56.000+09:00
このサイトは(株)国際文献社によって運用されています。