はやぶさ2が小惑星リュウグウから持ち帰ったサンプルには,鏡像異性体のL型とD型のアミノ酸が同じ比率で含まれていた.一方,地球上の生物がタンパク質合成に使うアミノ酸はL型のみであることから,D型アミノ酸は,原核生物での細胞壁合成などに使われる例外的存在と考えられてきた.TCA回路の発見者であるKrebsは,1930年代に肝臓と腎臓でD-アミノ酸の酸化酵素を発見したが,当時は基質となるD-アミノ酸は天然には存在しないと考えられていた.キラルアミノ酸や微量アミノ酸の分離分析技術の発達により,哺乳類脳でのD-セリン(D-Ser)の発見やD-Serを合成するセリン異性化酵素(serine racemase:SRR)の発見が新たな研究分野を拓き,現在ではD-Ser,D-Asp,D-Alaなどが存在することが明確となっている.2003年に私は富山医科薬科大学医学部で分子神経科学講座を主宰する機会に恵まれ,それまで行ってきたN-methyl-D-aspartate(NMDA)型グルタミン酸受容体(NMDAR)の分子生物学的研究から,その活性を制御しているD-Serに着目し研究を始めた.本稿では,我々の研究を中心としてD-Serによる生体機能調節について紹介する.
NMDARは,興奮性神経伝達,シナプス可塑性,神経回路形成,高次脳機能,脳の病態などに関わる重要な陽イオン選択的チャネル分子である1).NMDARチャネルの活性化のためには,アゴニストとしてGluやNMDAが結合するのみならず,コ・アゴニスト(アゴニストとともに作用して受容体を十分に活性化させる物質)としてGly,D-Ser,D-Alaなどが必要であることが示された2).脳内にGlyは豊富に存在するが,通常はGlyトランスポーターの取り込みによりシナプス間隙のGly濃度は低いと考えられること3),リコンビナントあるいは脳神経細胞上のNMDARに対する親和性はD-Serの方がGlyより高いこと4, 5),GlyではなくD-Serが大脳皮質視覚野や海馬由来脳切片でのNMDARによる神経伝達や,NMDAR依存的なシナプス伝達の長期増強(long-term potentiation:LTP)の誘導に関わること6, 7),などからD-SerはNMDARの内在性コ・アゴニストと考えられている.
D-Serが1992年にラット脳内に発見されて8)以来,その合成,分解,輸送などの動態制御の分子機構に注目が集まっていた.D-Serの合成に関しては,ラット脳由来シナプトソームに,L-phosphoserineを加えるとD-Serが増加することからphosphoserine phosphatase(PSP)の関与の可能性9)や,Gly開裂系(glycine cleavage system:GCS)に障害がある非ケトン性高Gly血症患者の死後脳でD-Serが減少する,あるいは,ラットへのGCS阻害薬の投与で脳内D-Serが減少する10)ことからGCSの関与の可能性が示唆されていた.後述するように1999年にセリン異性化酵素(SRR)が発見され,SRRの遺伝子欠損マウスの解析から前脳のD-Serの90%程度は主にSRRにより合成されることが明らかとなった.
D-アミノ酸の分解については,D-アミノ酸酸化酵素(D-amino acid oxidase:DAAO)がTCA回路の発見者であるKrebsによって1930年代にブタの肝臓と腎臓で発見された11)が,当時は自然界に基質となるD-アミノ酸は存在しないと考えられていた.その後,哺乳類ではD-Ser,D-Asp,D-Alaなどが発見され,これらのうち中性アミノ酸のD-SerとD-AlaはDAAOが分解を行う一方,酸性アミノ酸であるD-AspはStillらが最初にウサギ腎臓で発見したD-Asp酸化酵素(D-Asp oxidase:DDO)12)がその分解に関わると考えられている.
D-Serの細胞間輸送には,中性アミノ酸トランスポーターが関わると考えられている.特に神経細胞に発現するAsc-1(Slc7a10)がD-Serの細胞外への放出に13, 14),System Aアミノ酸トランスポーター(Slc38a1, Slc38a2)が神経細胞への取り込みに関わり15),アストロサイトに発現するアミノ酸交換輸送体ASCT1(Slc1a4)がD-Serの取り込みに主に関与している16)と考えられている.アストロサイトと神経細胞間のL-SerとD-Serの輸送に関しては,セリンシャトル仮説17)が提唱され,この仮説を支持する研究結果も報告されている18).
1999年にD-Serを合成するセリン異性化酵素(SRR)が,Woloskerらによりラット脳より精製され19),さらにcDNAクローニングされそのアミノ酸配列一次構造が明らかになった20).SRRは,pyridoxal 5′-phosphate(PLP)依存的酵素で,L-SerとD-Serの異性化反応を触媒する20)とともに,Serの脱水反応によりピルビン酸とアンモニアを生成する脱水酵素活性21)も有している.これらの酵素反応機構の詳細などについては,いくつかの総説がある22, 23).遺伝子クローニングされ,リコンビナント酵素で検討されたSRRの活性は個体でのD-Serの存在量を説明できるのか不明であった.そこで,我々を含むいくつかのグループがSRRの遺伝子ノックアウト(SRR-KO)マウスを作製して解析を行った.その結果,SRR-KOマウスの大脳皮質や海馬では,野生型マウスに比べD-Ser量が10~20%程度にまで低下し,SRRが脳内での主要なD-Ser合成酵素遺伝子であることが証明された24–26).
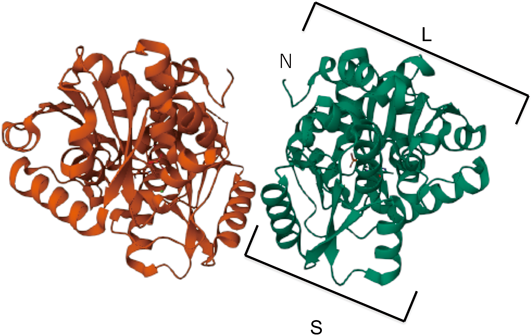
SRRはNMDARの内在性コ・アゴニストのD-Serを合成することや,SRR-KOマウスがアルツハイマー病(AD)の原因と考えられるアミロイドβペプチドの脳内急性投与による神経細胞死の誘導に対して抵抗性を示すこと24)から,SRRはNMDARの過剰活性化による神経変性疾患の治療薬標的になる可能性が高いと考えられ,その立体構造の解析が進められた.ヒトとラットの変異型SRRの結晶立体構造がSmithらにより初めて報告された27).SRRは大小の二つのドメインからなる対称型二量体を形成して機能すると考えられている.報告されたSRRの立体構造情報27),ならびに我々が明らかにしたリコンビナント野生型ヒトSRRの立体結晶構造28)(PDB 5X2L;図1)を基としてin silicoスクリーニング,in vitroでのSRR活性阻害効果の検証を行い,いくつかの新たなSRR阻害薬候補を創製し報告した28–30).SRRの立体構造を基盤としたin silicoスクリーニングで得られた候補化合物の多くは疎水性が高く,水溶液の酵素活性測定系では扱いが難しく,現時点でも満足できるSRR阻害薬は得られていない.また,創薬を進める際には個体レベルでの薬物作用を示さないと企業などとの共同研究に発展できないことも経験した.
SRRの活性調節機構として,Mn2+,Mg2+,Ca2+などの2価イオンやATPなどの小分子で増強されること,また,glutamate receptor interacting protein(GRIP)やprotein interacting with C-kinase(PICK1)はSRRと結合し,その活性増強に関わること,Golga-3やdisrupted in schizophrenia 1(DISC1),FBXO22などとの結合はSRRのタンパク質としての安定化制御に関わることなどが報告され,総説22, 23)としてまとめられている.このようにSRRは細胞内や個体内ではさまざまな分子との相互作用により活性調節を受けている.
SRR機能に重要なアミノ酸配列がアミノ酸変異導入等の解析により明らかにされている(図2).特に,酵素活性に必須なのはPLPが結合する56番目のLysであり,SRR-K56G変異体は活性がなくなる20, 31).また,基質であるSerの異性化反応と脱水反応には基質の保持に関わる84番目のSerが反応中間体の形成に重要である22, 23).脱水反応には152~155番目のHis–Pro–Asn–Glnの配列が関わっており,特に155番目のGlnをAspに置換したSRR-Q155Dでは,異性化反応が促進される一方,脱水反応が大きく抑制され,培養細胞に過剰発現させると細胞内にD-Serが蓄積する21, 31).また,152~154番目のHis–Pro–Asnの配列は,基質としてのSerの認識に重要である.SRRには弱いながらもアスパラギン酸異性化酵素(Asp racemase:DR)の活性がある32, 33).進化の過程でSRRを祖先形として遺伝子重複でDRが生じたと考えられているが,DRでは,同じ位置にSerを有している.たとえば,SRRの152~154番目のHis–Pro–Asnの配列の3アミノ酸をSer–Ser–Serに置換する(triple Ser構造)と,Aspを基質とするDRとして高い活性を示す34).
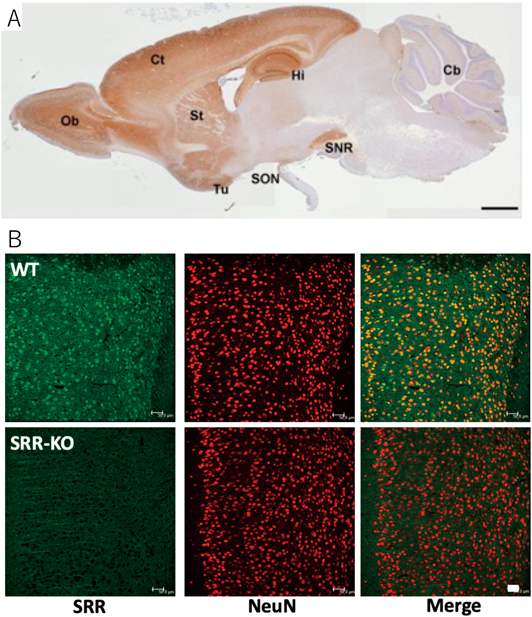
SRRの遺伝子がクローニングされた際,抗体を用いた免疫組織化学法によりその発現がアストロサイトに特異的とされ,SRRにより産生されるD-Serはグリア細胞から放出されて神経細胞の機能調節に関わる“グリオトランスミッター(Gliotransmitter)”の一つであると提唱された.しかしながら,新たな特異性の高い抗体とSRR-KOマウスを用いた検討などにより35, 36),SRRは通常は神経細胞に発現していると考えられた(図3).D-Serが神経細胞由来かアストロサイト由来かに関しては,それぞれの立場から議論が盛んに行われている37–39)が,正常な脳内ではSRRが神経細胞に発現し,D-Serは主に神経細胞に由来する一方,炎症の際には,後述するようにアストロサイトやミクログリアなどでもSRR発現が誘導されると考えられている.
我々を含むいくつかのグループがSRRの遺伝子ノックアウトマウスを作製した24–26, 36).我々のSRR-KOマウスはC57BL/6N系統由来の胚性幹(embryonic stem:ES)細胞を用いて標的遺伝子組換えにより作製した24, 36).我々の作製したSRR-KOマウスの大脳皮質,海馬では,D-Serが野生型コントロールマウスに比べて約10%程度であり,SRRが前脳の90%のD-Ser合成に関わる主要な酵素であることが明らかとなった24).同様の結果は他のグループのSRR-KOマウスでも得られており,D-Serの低下に伴ってSRR-KOマウスでは,後述するようにNMDARの機能低下が生じている.残りの10%程度の脳内D-Serの起源については,腸内細菌に由来することが想定されていたが,腸内細菌叢を除いても脳内D-Serは減少しないことから40),前述したPSPやGCSなどの関与を検証する必要がある.
記憶は,獲得,固定,想起,想起後の不安定化と再固定,消去などのそれぞれの過程でさまざまな制御を受けている.海馬依存的な連合記憶の一つである文脈依存恐怖条件づけの分子機構を明らかにすることは,異常な恐怖反応を伴う心的外傷後ストレス障害(post traumatic stress disorder:PTSD)の機構を明らかにし効果的な治療法を開発する上で重要である.マウスを用いた文脈依存恐怖条件づけでは,条件づけ箱の中で一定時間マウスに探索行動をさせた後,足に弱い電気ショックを非条件刺激(unconditioned stimulus:US)として与える.条件づけ箱の状況が文脈として条件刺激(conditioned stimulus:CS)となる.条件付けが成立するとマウスは,同じ条件づけ箱に入れられるとCSに対する条件反応(conditioned response:CR)としてすくみ応答(フリージング)を恐怖反応として示す.このフリージング時間を,記憶の指標として定量的に評価する.
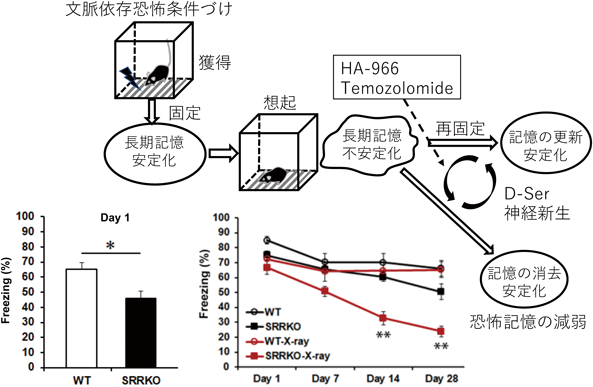
文脈依存恐怖条件づけの獲得には,海馬のNMDARが必要である.そこで,NMDARのコ・アゴニストのD-Serが低下したSRR-KOマウスを用いて,記憶能力を評価するために恐怖条件づけの行動を解析した.恐怖条件づけの記憶獲得にはSRR-KOマウスでは障害はみられなかったが,翌日の記憶想起テストでは,SRR-KOマウスは有意な記憶低下を示した(図4).マウスでは翌日に評価される記憶は長期記憶として扱う.したがってこの結果から,長期記憶の形成にD-Serが関わることが示された41).ヒトのPTSDに対する認知行動療法として,安全な状況下でトラウマ記憶を思い出す曝露療法があり,曝露時に想起されるトラウマ記憶が安全であることを新たに学習することで,トラウマ記憶を消去する方法として用いられている.そこで,SRR-KOマウスを用いて,さまざまな記憶消去法に対する反応を調べた.その結果,恐怖条件づけ直後(15分後)に30分間の消去トレーニング(30分間条件づけ箱に入れCSは同じであるが,USとしての電気ショックを与えない)を行うとSRR-KOマウスでは恐怖記憶の低下が野生型マウスより促進していた.一方,条件づけ翌日に消去トレーニングを行うと野生型マウスと差がなかったが,記憶想起2時間後から消去トレーニングを行うとSRR-KOマウスでは記憶消去効果がみられなかった41).記憶想起2時間後のSRR-KOマウスの海馬ではシナプス可塑性のLTPに関わるAMPA型グルタミン酸受容体(GluR)のGluA1サブユニットが増加することや,SRR-KOマウスで記憶想起の直前にD-Serを投与するとGluRA1の量が低下し記憶消去効果が観察された.これらの一連の研究の結果から,D-Serは記憶の固定と想起後の再固定に対する消去トレーニング過程で異なる機能を担っていることが明らかとなった41).我々の研究結果は,記憶想起後にNMDARの機能を増強することで,PTSDの異常な恐怖記憶に対する効果的な消去が可能であることを示唆している.
また,SRR-KOマウスの海馬では,ダブルコルチン(DCX)陽性の未熟神経細胞数が増加しており成体神経新生が増加していた.そこで,この神経新生の機能を明らかにするために,海馬での成体神経新生をX線照射により低下させたところ,文脈依存恐怖条件づけ課題で記憶想起を繰り返すことで,恐怖反応が効果的に減弱することを見いだした42)(図4).野生型マウスにNMDARのD-Ser結合サイトに対する阻害剤HA-966と細胞増殖阻害薬(Temozolomide:TMZ)の両方を投与することでも同様の恐怖反応の低下が観察され,恐怖反応の低下が薬物を用いても可能であった.これらの研究の結果から,記憶想起後の再固定化のステップに,D-SerによるNMDARの活性化と成体神経新生の両方が重要であることが明らかとなり,PTSDの新たな治療法の提案にもつながる可能性がある(図4).
1)統合失調症との関係
SRR-KOマウスでは,NMDARの機能低下が起きているため,統合失調症の発症仮説に対応したモデルとなる可能性がある.統合失調症は,幻覚や妄想などの陽性症状,自閉や感情の平板化などの陰性症状,認知機能障害を特徴とする精神疾患である.我々のC57BL/6N系統を遺伝的背景とするSRR-KOマウスは,統合失調症患者ならびにモデル動物で観察される音驚愕反射のプレパルス抑制(prepulse inhibition:PPI)に異常は観察されなかったため43),それ以上の評価は行わなかった.一方,Coyleらのグループが作製したSRR-KOマウス25)は,PPIの異常は示さないが統合失調症の陽性症状の一つとして評価される新規環境での運動量の増加,モリス水迷路での記憶の障害があることからさまざまな検討が行われている.たとえば,一次体性感覚野の錐体神経細胞で樹状突起数やスパイン密度の低下,大脳皮質体積の減少,脳由来神経栄養因子(brain-derived neurotrophic factor:BDNF)の発現低下が示され44),統合失調症と同様の神経発達の障害が示唆された.さらに,海馬の体積減少と歯状回顆粒神経細胞での樹状突起スパイン密度の低下などの形態学的異常,歯状回でのLTP維持の障害やBDNFシグナルの減少などの生化学的異常,7回の電気ショックを与えて形成される文脈依存恐怖条件づけと,扁桃体が関わる音手がかり刺激依存恐怖条件づけの両方の障害の認知行動学的異常も報告されている45).これらの生化学的障害と行動学的障害は,D-Serを20日間連続投与することで正常化できる45).これらのことから,SRR-KOマウスの統合失調症モデルとしての有用性と,その治療への慢性的なD-Ser投与の有用性が提唱されている.このSRR-KOマウスでは,統合失調症の陰性症状や社会性障害の有無も検討され,行動課題では軽微ながら有意な社会性の障害,超音波コミュニケーションの異常,新規物体認知機能の異常が報告されている46)が,これらの障害に対するD-Ser投与の効果は検討されていない.また,核磁気共鳴イメージング(magnetic resonance imaging:MRI)を用いた研究で,SRR-KOマウスの脳室の拡大,GluとGABA含量の増大が見いだされている47).GluとGABAの上昇については,統合失調症でのNMDAR機能低下に対する代償性のGluの上昇や,発火頻度の高いパルブアルブミン陽性の抑制性神経細胞の機能低下に対する代償的なGABA含有量の増加の可能性が考察されている.
統合失調症は,NMDARの機能低下とともにドパミン神経伝達過剰が生じていると考えられ,臨床では特にドパミンD2受容体に対する阻害薬が陽性症状の治療に用いられている.CoyleらのSRR-KOマウスは,前頭皮質脳切片での電気生理学と薬物を用いた解析から,ドパミンのD1受容体とD3受容体の活性化に伴うNMDARを介した神経伝達とシナプス可塑性の調節に異常があることが示された48).さらに前頭皮質の機能が関わる新規物体認知試験でのSRR-KOマウスの成績の低下が,D-Serの投与で正常化された.また,SRRの遺伝子プロモーター部位の変異が遺伝子発現に影響し,ヒトの統合失調症のリスク遺伝子であることも報告されている49).
ウィルス感染等がきっかけとなり発症する抗NMDAR自己抗体脳炎は,初期には統合失調症様の行動異常を示し,その後てんかん発作や呼吸抑制などの重篤な症状を引き起こす50, 51).統合失調症にも抗NMDAR自己抗体によるNMDARの機能低下が関わる可能性がある.我々も,統合失調者症患者で自己抗体の存在を検索し自己抗体価の高い患者を見いだし,D-Serを投与することで症状が軽快することを報告している52).以上の結果から,SRR-KOマウスはNMDAR機能低下による統合失調症モデルとして解析を行う価値が高いこと,また統合失調症には原因となる背景が異なる集団がいくつか存在する可能性があり,これらの集団を区別してD-Serの治療効果を検証する必要があると考えられる.
2)薬物依存との関係
NMDARは依存薬物投与による行動異常にも関与している.精神刺激薬のメタンフェタミン(METH)の急性投与で誘発される行動量の上昇は,我々の作製したSRR-KOマウスでも野生型コントロールマウスと同様に観察される.一方,METH連続投与後の低容量のMETHで誘発される行動量の上昇(行動の鋭敏化)が,SRR-KOマウスでは観察されなかった53).また,SRR-KOマウスでは,METH投与により誘導される側坐核でのドパミン遊離量が低下しており,METH投与後の神経活動活性化に対応するシグナル伝達を反映するERK1/2分子のリン酸化の上昇もSRR-KOマウスの線条体では観察されなかった.これらの結果から,薬物依存の形成にD-Serが関わる可能性が示唆された.
3)うつ病との関係
うつ病にもNMDARが関わっていると考えられている.たとえばNMDARのチャネルブロッカーであるケタミン(ketamine)を低容量投与することが,大うつ病に対して即効的で持続的な改善効果を引き起こす54).また,うつ病患者の脳脊髄液のD-Ser濃度とうつ症状には逆相関の関係が報告されている55).我々のSRR-KOマウスでは,野生型コントロールマウスで観察される社会性敗北ストレスで誘発されるうつ様症状がほとんど観察されず,抵抗性を示した56).SRR-KOマウスでは,野生型コントロールマウスでうつ症状に付随する脳内BDNFの発現変化やBDNF受容体のTrkBのリン酸化などの生化学的変化も起きていなかった.興味深いことに,SRR-KOマウスにD-Serを飲水から摂取させると,社会性敗北ストレスで誘発されるうつ様症状を示した56).したがって,D-Serを減少させNMDARの機能低下が引き起こされるとストレスに対する抵抗性を高め,うつのリスクを減らす可能性が示唆された.
1)アルツハイマー病との関係
我々が作製したSRR-KOでは,脳内D-Serが野生型マウスの10%程度にまで低下していたためNMDARの機能低下が想定された.NMDARの機能を評価するために,脳内にNMDAを急性投与したところ,誘発される神経細胞死が,野生型コントロールマウスに比べてSRR-KOマウスでは軽減していた24).さらに,神経毒性を示しアルツハイマー病(AD)の原因と考えられるアミロイドβ(Aβ)ペプチドの海馬内急性投与による神経細胞死も軽減していた.ADを含む多くの変性疾患での神経細胞死にはNMDARを介した興奮毒性が関わっていると考えられており,NMDARチャネルブロッカーの一つのメマンチン(memantine)がADの治療薬として使用されている.またADの遺伝学的モデルマウスでSRRの遺伝子ノックアウトを行うと神経細胞死が軽減することも観察している57).我々の研究結果からADではNMDARの機能を低下させること,つまりD-Ser濃度を下げることがADに対する治療につながると考えられる.一方で,ADのモデル動物にD-SerやL-Serを投与することで認知機能の低下が抑制され,AD進行を遅らせるとの報告もある58).これらの結果は,AD進行過程の異なるタイミングを評価している可能性があり,慎重に検討する必要がある59).また,AD患者ではSRRの発現が炎症性のA1アストロサイトで検出されている60)との報告もあり,炎症を抑えることが異常なD-Serの機能抑制やADの治療につながる可能性がある.
2)網膜変性との関係
神経変性モデルの一つとして,糖尿病性網膜症に伴う視神経細胞死がある.マウス網膜では,SRRは顆粒細胞層に発現している61).1型糖尿病動物モデルとして膵臓β細胞を障害するストレプトゾシン(streptozocine:STZ)投与モデルがあるが,野生型コントロールマウスおよびSRR-KOマウスにSTZを投与すると,1週間後から両群のマウスとも血糖の上昇が起こり糖尿病マウスとなる.野生型コントロールマウスではSTZ投与6か月後に網膜を解析するとSRRの発現が顆粒細胞層で上昇している傾向を認めた.さらに,網膜神経節細胞のマーカーであるBrn3陽性細胞数が糖尿病マウスで減少し神経細胞死が起きていることが示唆された.一方,SRR-KOマウスでは野生型コントロールマウスに比べ障害の程度が低くBrn3陽性細胞数もより多く残っており,また内網膜層の障害を示す菲薄化の程度も軽減していた61).さらに糖尿病性網膜症で観察される無細胞性の異常な毛細血管の数もSRR-KOでは少なかった.ラットにSTZを投与して作製された糖尿病性網膜症モデルでもSRRの網膜での発現上昇と眼球内の房水でのGluとD-Serの上昇が報告されている62).これらの結果から,糖尿病性網膜症に対してSRRの活性を抑え,D-Serを減少させることが保護的治療につながる可能性があることが示唆された.
3)てんかんとの関係
NMDARの過剰活性化はてんかん発作を引き起こす.過剰なてんかん発作はさらに興奮毒性による神経細胞死にもつながる可能性がある.脳全体に広がる全般性てんかん発作モデルの一つとしてGABA受容体阻害薬のペンチレンテトラゾール(pentylenetetrazole:PTZ)投与による薬理学的モデルがある.PTZは投与量によりさまざまなステージの全般性てんかん発作を引き起こす.発作の程度は行動学的指標により解析され,発作発現までの時間,発作ステージ,発作の継続時間などで評価することができる.SRR-KOマウスは,野生型コントロールマウスに比べてPTZ誘発による全般性発作の継続時間の短縮や最も重篤なステージ4に至る個体がいないなど,発作抵抗性を示した63).また,大脳皮質,海馬などで神経活動に依存して発現するc-FOSの陽性細胞が野生型マウスに比べて少なかった.さらに,SRR-KOマウスでは海馬歯状回での細胞外Glu濃度が低く,野生型コントロールマウスでPTZ投与30分後と150分後に検出されたGluの濃度上昇変化も観察されなかった.これらの結果からD-Serがプレシナプスに存在するNMDARを介してGlu放出の制御に関わる可能性が示唆されるとともに,SRRはてんかん発作を軽減する薬物標的となると考えられた.
その他の神経変性を伴う病態として外傷性脳障害(traumatic brain injury:TBI)がある.マウスTBIモデルでは,障害後に炎症に関わるアストロサイトやミクログリアでSRRの発現が上昇し,D-Serの産生と放出が増強することでシナプス伝達や可塑性の障害が生じると提唱されている64, 65).一方で,マウスTBIモデルでD-Serを増加させると考えられるDAAOの阻害薬(6-chlorobenzo[d]isoxazol-3-ol:CBIO)の投与が障害の程度を軽減し,BDNFの発現を促進して,神経保護的作用を示す可能性も報告されている66, 67).これらの一見矛盾するD-Serの役割の違いは,TBIモデルの違いや,TBIの進行過程の違いにより,シナプス障害に伴う認知機能障害にはD-Serの補充による保護的効果がある一方,さらに障害が進行した炎症過程では,グリア細胞由来の制御されないD-Serの放出が障害の拡大に関わっている可能性がある.
4)疼痛との関係
NMDARは痛覚伝導にも関わることから,我々のSRR-KOマウスを用いて,後肢足裏へのホルマリン投与による急性炎症痛モデルを用いて痛覚伝導を評価した.その結果,SRR-KOマウスは野生型コントロールマウスに比べて,疼痛行動の上昇を示し,痛覚過敏となっていることが明らかとなった68).痛覚過敏に伴いSRR-KOマウスでは,神経伝達の上昇を示す脊髄後角でのc-FOS発現細胞数の増加や細胞内シグナル伝達の増強を示唆するERK1/2のリン酸化の上昇も認めた.野生型コントロールマウスでは,SRRは脊髄後角の感覚神経細胞に主に発現し,ホルマリン投与を行うと発現が減少するとともに,その細胞内分布が変化することが観察された.SRRは細胞内での分布変化により細胞膜に結合して不活性化される機構が培養細胞で報告されている69)ことから,野生型コントロールマウス脊髄での急性疼痛によるSRRの発現減少や,細胞内分布の変化は,疼痛を抑制するための機構である可能性が示唆された.一方で,関節炎モデルのような慢性炎症疼痛では,SRR-KOマウスでは痛みに伴う超音波鳴声数が上昇しないことも報告されているので70),痛みの種類や痛覚伝導路によってD-Serの役割が異なっている可能性がある.
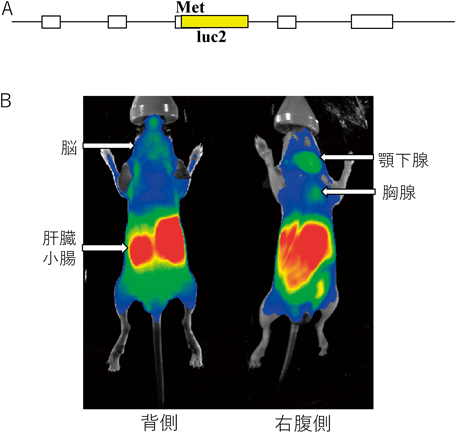
我々のSRR-KOマウスでは,腎臓,筋肉,精巣でD-Serの減少が観察されたが,肝臓,膵臓,前立腺,心臓,肺,眼球などではD-Serの濃度は野生型コントロールマウスと変化はなかった71).SRRの全身での発現を検討するためにSRRの遺伝子座にホタルの発光遺伝子(luc)を導入したトランスジェニックマウスを作製したところ,SRRの発現を示す発光シグナルは末梢臓器にも観察された(図5).これらの臓器でSRRはさまざまな生理機能と病態に関わっていると考えられる.
1)腎臓におけるD-セリンの役割
腎臓にはD-Serの合成に関わるSRR,分解酵素のDAAO, NMDARが発現している.NMDARチャネルブロッカーの(+)-MK-801やD-Serのラット腎臓への投与が,濃度依存的に尿流量やNa+排泄量を減少させ,また求心性の腎神経活性を上昇させることから腎機能の調節に関わっていると考えられている72).また腎臓ではL-SerとD-Serは排出されたのち近位尿細管から再吸収されるが,近位尿細管にはDAAOが発現しD-Serの分解を行っている73).その際,過剰量のD-Ser投与はDAAOによる分解で生じるH2O2による腎毒性を引き起こすと考えられている74).一方,腎障害ではD-Serの分解が障害され血漿中で上昇するため,D-Serは急性腎障害や慢性腎障害の有用なバイオマーカーとして腎機能の評価に使うことが可能である75, 76).興味深いことに腎臓のD-Serには腸内細菌由来のものも含まれ,急性腎障害でのD-Serの上昇が腎臓に対する保護作用を示すと考えられている76).
2)表皮におけるD-セリンの役割
以前より皮膚にNMDARが発現していることが報告されていることから,我々はSRRやD-Serが皮膚に存在しているか調べた.その結果,皮膚分化の最終段階の顆粒細胞層や脱核した角質層でSRRが発現していることを見いだした77).また培養ケラチノサイトにはD-Serの合成能が認められた.そこで皮膚でのSRRの機能を明らかにするためにSRR-KOマウスの皮膚を解析した.その結果,SRR-KOマウスは形態学的に皮膚の角質層が有意に薄く,顆粒細胞層でのケラトヒアリン顆粒の蓄積大型化が観察され,皮膚の角化の障害が示唆された.実際にSRR-KOマウスでは皮膚からの水分蒸発量が大きく,角層除去後の回復も遅いことから皮膚バリア機能が低下していることが明らかとなった77).さらに皮膚バリア機能不全を示すアトピー性皮膚炎の患者表皮では,SRRの発現が上昇するとともに,含有D-Ser濃度が上昇していた.ヒト培養ケラチノサイトでは,炎症性サイトカインであるtumor necrosis factor(TNF)-αあるいはmacrophage migration inhibitory factor(MIF)の投与によりSRRの発現が誘導された78).これらの結果から,皮膚においてその生理機能と病態にSRRおよびD-Serが関わっていることが明らかとなった.
3)その他の臓器におけるD-セリンの役割
その他の末梢臓器では,SRRの発現は肝臓に検出されているが71),発現細胞や明確な機能についての情報は現時点ではない.末梢臓器でのD-Serの役割についてまとめた総説79)があるが,培養細胞での解析結果など個体での機能を十分に反映していない可能性があり,今後の解析が必要である.
D-Serはさまざまな生物に存在している.植物ではD-Serは増殖を抑制すると考えられてきた.しかしながら,シロイヌナズナ(Arabidopsis thaliana)ではD-SerがNMDAR相同分子を活性化しCa2+の流入を介して花粉管の伸長を促すことが報告されている.これら植物でのD-Serの機能に関しては,SRRとDRの進化の観点からの総説がある80).
これまで主に哺乳類でのD-SerやSRRの機能を述べてきた.無脊椎動物でのSRRとDRの分布と進化については総説がある33).個別の無脊椎動物種でD-SerやSRRの生体機能が明らかになっているものはそれほど多くはない.線虫(Caenorrhabditis elegans)では,セリンラセマーゼ遺伝子(Serr-1)は,腸管と皮下組織に発現し,飢餓状態で速やかにD-Serを合成する.Serr-1遺伝子欠損線虫では,長期飢餓状態での探餌の際の適応的な運動が障害されている81).ショウジョウバエ(Drosophila)では,セリンラセマーゼ(sr)遺伝子は,腹神経束(ventral nerve cord)と中腸の腸細胞(enterocyte)に発現している.Sr遺伝子欠損ハエは,夜間の睡眠時間の有意な減少が観察され,腸管でSr遺伝子を発現させることで睡眠の障害から回復する82).また,D-Serはホヤ(Ciona intestinalis type A)の変態過程に関与している.ホヤのセリンラセマーゼ遺伝子(Ci-SRR3)を破壊すると遊泳体から固着体への変態過程で尾部の吸収に関わる皮膚細胞内の小胞の分泌が障害され,尾部の吸収不全が引き起こされる.NMDAR相同分子もホヤに存在することから,SRRによって作られるD-SerがNMDARに作用し皮膚細胞からの小胞分泌を制御していると考えられる83).この障害は,SRR-KOマウスの皮膚で観察されたケラトヒアリン顆粒の蓄積大型化77)と似ており,進化的に保存されたD-Ser機能の一端を示している可能性がある.
SRRにはD-Ser以外にD-Aspの合成能力がある32, 33).また,SRR-KOマウスの海馬ではD-Aspの濃度が低下している84)が,哺乳類ではD-Aspの合成酵素は見つかっておらず,SRRが個体レベルでD-Asp合成にどの程度関わっているのかは現時点で不明である.また胎仔マウスでD-Cysが存在することが報告されたが,SRR-KOマウスではD-Cysの濃度も低下しているため,D-Cysの合成にもSRRは関与すると考えられる85)が,その経路は不明である.D-アミノ酸もこれまで知られているL-アミノ酸の代謝酵素で代謝されるのか,またその際の選択性や効率はどの程度なのかを明らかにすることで,D-アミノ酸の生体内代謝の全貌が明らかになると考えられる.最初にも述べたように,D-アミノ酸の体内動態には合成酵素のみならず代謝酵素,分解酵素,輸送体が関わることから,これらの役割についても検証する必要がある.
また,SRRは生産するD-Serを介してNMDARの機能に関わりさまざまな病態にも関わることが明らかになってきたので,SRRを標的とした活性化薬や阻害薬あるいは遺伝子発現制御薬の開発がNMDARの機能低下による精神疾患や認知障害,機能過剰による神経変性疾患に新たな治療法を提供できる可能性がある.しかしながら,述べてきたようにSRR-KOやD-Ser操作の効果が必ずしも一致していない場合がある.病態進行過程や症状においてD-SerやSRRが機能低下しているのか,機能過剰になっているのか,どの細胞や臓器で変化が起きているのかを正しく評価して介入することが,疾患に対するよりよい治療法の開発につながると考えられる.
謝辞Acknowledgments
本稿をまとめるに際し,これまで一緒に多くの研究を担ってくれた富山大学医学部分子神経講座のスタッフ,大学院生,学部学生に感謝いたします.また縁あって多くの研究成果にご協力いただきました共同研究者各位にも感謝いたします.
引用文献References
1) Mori, H. (2017) Overview of the NMDA receptor. in The NMDA Receptor (Hashimoto, K. ed.), The Receptor. Vol. 30, pp. 1–18, Humana Press.
2) Kleckner, N.W. & Dingledine, R. (1988) Requirement for glycine in activation of NMDA-receptors expressed in Xenopus oocytes. Science, 241, 835–837.
3) Papouin, T., Ladépêche, L., Ruel, J., Sacchi, S., Labasque, M., Hanini, M., Groc, L., Pollegioni, L., Mothet, J.P., & Oliet, S.H. (2012) Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell, 150, 633–646.
4) Matsui, T., Sekiguchi, M., Hashimoto, A., Tomita, U., Nishikawa, T., & Wada, K. (1995) Functional comparison of D-serine and glycine in rodents: the effect on cloned NMDA receptors and the extracellular concentration. J. Neurochem., 65, 454–458.
5) Berger, A.J., Dieudonné, S., & Ascher, P. (1998) Glycine uptake governs glycine site occupancy at NMDA receptors of excitatory synapses. J. Neurophysiol., 80, 3336–3340.
6) Henneberger, C., Papouin, T., Oliet, S.H., & Rusakov, D.A. (2010) Long-term potentiation depends on release of D-serine from astrocytes. Nature, 463, 232–236.
7) Meunier, C.N.J., Dallérac, G., Le Roux, N., Sacchi, S., Levasseur, G., Amar, M., Pollegioni, L., Mothet, J.-P., & Fossier, P. (2016) D-Serine and glycine differentially control neurotransmission during visual cortex critical period. PLoS One, 11, e0151233.
8) Hashimoto, A., Nishikawa, T., Hayashi, T., Fujii, N., Harada, K., Oka, T., & Takahashi, K. (1992) The presence of free D-serine in rat brain. FEBS Lett., 296, 33–36.
9) Wood, P.L., Hawkinson, J.E., & Goodnough, D.B. (1996) Formation of D-serine from L-phosphoserine in brain synaptosomes. J. Neurochem., 67, 1485–1490.
10) Iwama, H., Takahashi, K., Kure, S., Hayashi, F., Narisawa, K., Tada, K., Mizoguchi, M., Takashima, S., Tomita, U., & Nishikawa, T. (1997) Depletion of cerebral D-serine in non-ketotic hyperglycinemia: possible involvement of glycine cleavage system in control of endogenous D-serine. Biochem. Biophys. Res. Commun., 231, 793–796.
11) Krebs, H.A. (1935) Metabolism of amino-acids: deamination of amino-acids. Biochem. J., 29, 1620–1644.
12) Still, J.L., Buell, M.V., Knox, W.E., & Green, D.E. (1949) Studies on the cyclophorase system; D-aspartic oxidase. J. Biol. Chem., 179, 831–837.
13) Rosenberg, D., Artoul, S., Segal, A.C., Kolodney, G., Radzishevsky, I., Dikopoltsev, E., Foltyn, V.N., Inoue, R., Mori, H., Billard, J.M., et al. (2013) Neuronal D-serine and glycine release via the Asc-1 transporter regulates NMDA receptor-dependent synaptic activity. J. Neurosci., 33, 3533–3544.
14) Sason, H., Billard, J.M., Smith, G.P., Safory, H., Neame, S., Kaplan, E., Rosenberg, D., Zubedat, S., Foltyn, V.N., Christoffersen, C.T., et al. (2017) Asc-1 transporter regulation of synaptic activity via the tonic release of D-serine in the forebrain. Cereb. Cortex, 27, 1573–1587.
15) Bodner, O., Radzishevsky, I., Foltyn, V.N., Touitou, A., Valenta, A.C., Rangel, I.F., Panizzutti, R., Kennedy, R.T., Billard, J.M., & Wolosker, H. (2020) D-Serine signaling and NMDAR-mediated synaptic plasticity are regulated by system A-type of glutamine/D-serine dual transporters. J. Neurosci., 40, 6489–6502.
16) Kaplan, E., Zubedat, S., Radzishevsky, I., Valenta, A.C., Rechnitz, O., Sason, H., Sajrawi, C., Bodner, O., Konno, K., Esaki, K., et al. (2018) ASCT1 (Slc1a4) transporter is a physiologic regulator of brain D-serine and neurodevelopment. Proc. Natl. Acad. Sci. USA, 115, 9628–9633.
17) Wolosker, H. (2011) Serine racemase and the serine shuttle between neurons and astrocytes. Biochim. Biophys. Acta, 1814, 1558–1566.
18) Neame, S., Safory, H., Radzishevsky, I., Touitou, A., Marchesani, F., Marchetti, M., Kellner, S., Berlin, S., Foltyn, V.N., Engelender, S., et al. (2019) The NMDA receptor activation by D-serine and glycine is controlled by an astrocytic Phgdh-dependent serine shuttle. Proc. Natl. Acad. Sci. USA, 116, 20736–20742.
19) Wolosker, H., Sheth, K.N., Takahashi, M., Mothet, J.-P., Brady, R.O. Jr., Ferris, C.D., & Snyder, S.H. (1999a) Purification of serine racemase: biosynthesis of the neuromodulator D-serine. Proc. Natl. Acad. Sci. USA, 96, 721–725.
20) Wolosker, H., Blackshaw, S., & Snyder, S.H. (1999b) Serine racemase: a glial enzyme synthesizing D-serine to regulate glutamate-N-methyl-D-aspartate neurotransmission. Proc. Natl. Acad. Sci. USA, 96, 13409–13414.
21) Foltyn, V.N., Bendikov, I., De Miranda, J., Panizzutti, R., Dumin, E., Shleper, M., Li, P., Toney, M.D., Kartvelishvily, E., & Wolosker, H. (2005) Serine racemase modulates intracellular D-serine levels through an α,β-elimination activity. J. Biol. Chem., 280, 1754–1763.
22) Raboni, S., Marchetti, M., Faggiano, S., Campanini, B., Bruno, S., Marchesani, F., Margiotta, M., & Mozzarelli, A. (2019) The energy landscape of human serine racemase. Front. Mol. Biosci., 5, 112.
23) Graham, D.L., Beio, M.L., Nelson, D.L., & Berkowitz, D.B. (2019) Human serine racemase: Key residues/active site motifs and their relation to enzyme function. Front. Mol. Biosci., 6, 8.
24) Inoue, R., Hashimoto, K., Harai, T., & Mori, H. (2008) NMDA- and β-amyloid1–42-induced neurotoxicity is attenuated in serine racemase knock-out mice. J. Neurosci., 28, 14486–14491.
25) Basu, A.C., Tsai, G.E., Ma, C.L., Ehmsen, J.T., Mustafa, A.K., Han, L., Jiang, Z.I., Benneyworth, M.A., Froimowitz, M.P., Lange, N., et al. (2009) Targeted disruption of serine racemase affects glutamatergic neurotransmission and behavior. Mol. Psychiatry, 14, 719–727.
26) Labrie, V., Fukumura, R., Rastogi, A., Fick, L.J., Wang, W., Boutros, P.C., Kennedy, J.L., Semeralul, M.O., Lee, F.H., Baker, G.B., et al. (2009) Serine racemase is associated with schizophrenia susceptibility in humans and in a mouse model. Hum. Mol. Genet., 18, 3227–3243.
27) Smith, M.A., Mack, V., Ebneth, A., Moraes, I., Felicetti, B., Wood, M., Schonfeld, D., Mather, O., Cesura, A., & Barker, J. (2010) The structure of mammalian serine racemase: evidence for conformational changes upon inhibitor binding. J. Biol. Chem., 285, 12873–12881.
28) Takahara, S., Nakagawa, K., Uchiyama, T., Yoshida, T., Matsumoto, K., Kawasumi, Y., Mizuguchi, M., Obita, T., Watanabe, Y., Hayakawa, D., et al. (2018) Design, synthesis, and evaluation of novel inhibitors for wild-type human serine racemase. Bioorg. Med. Chem. Lett., 28, 441–445.
29) Mori, H., Wada, R., Li, J., Ishimoto, T., Mizuguchi, M., Obita, T., Gouda, H., Hirono, S., & Toyooka, N. (2014) In silico and pharmacological screenings identify novel serine racemase inhibitors. Bioorg. Med. Chem. Lett., 24, 3732–3735.
30) Mori, H., Wada, R., Takahara, S., Horino, Y., Izumi, H., Ishimoto, T., Yoshida, T., Mizuguchi, M., Obita, T., Gouda, H., et al. (2017) A novel serine racemase inhibitor suppresses neuronal over-activation in vivo. Bioorg. Med. Chem., 25, 3736–3745.
31) Talukdar, G., Inoue, R., Yoshida, T., Ishimoto, T., Yaku, K., Nakagawa, T., & Mori, H. (2017) Novel role of serine racemase in anti-apoptosis and metabolism. Biochim. Biophys. Acta, Gen. Subj., 1861(1 Pt A), 3378–3387.
32) Ito, T., Hayashida, M., Kobayashi, S., Muto, N., Hayashi, A., Yoshimura, T., & Mori, H. (2016) Serine racemase is involved in D-aspartate biosynthesis. J. Biochem., 160, 345–353.
33) Uda, K., Abe, K., Dehara, Y., Mizobata, K., Sogawa, N., Akagi, Y., Saigan, M., Radkov, A.D., & Moe, L.A. (2016) Distribution and evolution of the serine/aspartate racemase family in invertebrates. Amino Acids, 48, 387–402.
34) Uda, K., Abe, K., Dehara, Y., Mizobata, K., Edashige, Y., Nishimura, R., Radkov, A.D., & Moe, L.A. (2017) Triple serine loop region regulates the aspartate racemase activity of the serine/aspartate racemase family. Amino Acids, 49, 1743–1754.
35) Kartvelishvily, E., Shleper, M., Balan, L., Dumin, E., & Wolosker, H. (2006) Neuron-derived D-serine release provides a novel means to activate N-methyl-D-aspartate receptors. J. Biol. Chem., 281, 14151–14162.
36) Miya, K., Inoue, R., Takata, Y., Abe, M., Natsume, R., Sakimura, K., Hongou, K., Miyawaki, T., & Mori, H. (2008) Serine racemase is predominantly localized in neurons in mouse brain. J. Comp. Neurol., 510, 641–654.
37) Wolosker, H., Balu, D.T., & Coyle, J.T. (2016) The rise and fall of the D-serine-mediated gliotransmission hypothesis. Trends Neurosci., 39, 712–721.
38) Papouin, T., Henneberger, C., Rusakov, D.A., & Oliet, S.H.R. (2017) Astroglial versus neuronal D-serine: Fact Checking. Trends Neurosci., 40, 517–520.
39) Wolosker, H., Balu, D.T., & Coyle, J.T. (2017) Astroglial versus neuronal D-serine: Check your controls! Trends Neurosci., 40, 520–522.
40) Osaki, A., Aoyama, M., Mita, M., Hamase, K., Yasui, M., & Sasabe, J. (2023) Endogenous D-serine exists in the mammalian brain independent of synthesis by serine racemase. Biochem. Biophys. Res. Commun., 641, 186–191.
41) Inoue, R., Talukdar, G., Takao, K., Miyakawa, T., & Mori, H. (2018) Dissociated role of D-serine in extinction during consolidation vs. reconsolidation of context conditioned fear. Front. Mol. Neurosci., 11, 161.
42) Inoue, R., Ni, X., & Mori, H. (2023) Blockade of D-serine signaling and adult hippocampal neurogenesis attenuates remote contextual fear memory following multiple memory retrievals in male mice. Front. Neurosci., 16, 1030702.
43) Mori, H. & Inoue, R. (2010) Serine racemase knockout mice. Chem. Biodivers., 7, 1573–1578.
44) Balu, D.T., Basu, A.C., Corradi, J.P., Cacace, A.M., & Coyle, J.T. (2012) The NMDA receptor co-agonists, D-serine and glycine, regulate neuronal dendritic architecture in the somatosensory cortex. Neurobiol. Dis., 45, 671–682.
45) Balu, D.T., Li, Y., Puhl, M.D., Benneyworth, M.A., Basu, A.C., Takagi, S., Bolshakov, V.Y., & Coyle, J.T. (2013) Multiple risk pathways for schizophrenia converge in serine racemase knockout mice, a mouse model of NMDA receptor hypofunction. Proc. Natl. Acad. Sci. USA, 110, E2400–E2409.
46) Matveeva, T.M., Pisansky, M.T., Young, A., Miller, R.F., & Gewirtz, J.C. (2019) Sociality deficits in serine racemase knockout mice. Brain Behav., 9, e01383.
47) Puhl, M.D., Mintzopoulos, D., Jensen, J.E., Gillis, T.E., Konopaske, G.T., Kaufman, M.J., & Coyle, J.T. (2015) In vivo magnetic resonance studies reveal neuroanatomical and neurochemical abnormalities in the serine racemase knockout mouse model of schizophrenia. Neurobiol. Dis., 73, 269–274.
48) Dallérac, G., Li, X., Lecouflet, P., Morisot, N., Sacchi, S., Asselot, R., Pham, T.H., Potier, B., Watson, D.J.G., Schmidt, S., et al. (2021) Dopaminergic neuromodulation of prefrontal cortex activity requires the NMDA receptor coagonist D-serine. Proc. Natl. Acad. Sci. USA, 118, e2023750118.
49) Morita, Y., Ujike, H., Tanaka, Y., Otani, K., Kishimoto, M., Morio, A., Kotaka, T., Okahisa, Y., Matsushita, M., Morikawa, A., et al. (2007) A genetic variant of the serine racemase gene is associated with schizophrenia. Biol. Psychiatry, 61, 1200–1203.
50) Dalmau, J., Lancaster, E., Martinez-Hernandez, E., Rosenfeld, M.R., & Balice-Gordon, R. (2011) Clinical experience and laboratory investigations in patients with anti-NMDAR encephalitis. Lancet Neurol., 10, 63–74.
51) Miya, K., Takahashi, Y., & Mori, H. (2014) Anti-NMDAR autoimmune encephalitis. Brain Dev., 36, 645–652.
52) Heresco-Levy, U., Durrant, A.R., Ermilov, M., Javitt, D.C., Miya, K., & Mori, H. (2015) Clinical and electrophysiological effects of D-serine in a schizophrenic patient positive for anti N-methyl-D-aspartate receptor antibodies. Biol. Psychiatry, 77, e27–e29.
53) Horio, M., Kohno, M., Fujita, Y., Ishima, T., Inoue, R., Mori, H., & Hashimoto, K. (2012) Role of serine racemase in behavioral sensitization in mice after repeated administration of methamphetamine. PLoS One, 7, e35494.
54) Mori, H. (2020) Site of ketamine action on the NMDA receptor. in Ketamine: From Abused Drug to Rapid-Acting Antidepressant (Hashimoto, Ide, & Ikeda eds.), pp. 47–67, Springer.
55) Ishiwata, S., Hattori, K., Sasayama, D., Teraishi, T., Miyakawa, T., Yokota, Y., Matsumura, R., Nishikawa, T., & Kunugi, H. (2018) Cerebrospinal fluid D-serine concentrations in major depressive disorder negatively correlate with depression severity. J. Affect. Disord., 226, 155–162.
56) Dong, C., Zhang, J.-C., Ren, Q., Ma, M., Qu, Y., Zhang, K., Yao, W., Ishima, T., Mori, H., & Hashimoto, K. (2018) Deletion of serine racemase confers D-serine –dependent resilience to chronic social defeat stress. Neurochem. Int., 116, 43–51.
57) Ni, X., Inoue, R., Wu, Y., Yoshida, T., Yaku, K., Nakagawa, T., Saito, T., Saido, T.C., Takao, K., & Mori, H. (2023) Regional contributions of D-serine to Alzheimer’s disease pathology in male AppNL-G-F/NL-G-F mice. Front. Aging Neurosci., 15, 1211067.
58) Le Douce, J., Maugard, M., Veran, J., Matos, M., Jégo, P., Vigneron, P.A., Faivre, E., Toussay, X., Vandenberghe, M., Balbastre, Y., et al. (2020) Impairment of glycolysis-derived L-serine production in astrocytes contributes to cognitive deficits in Alzheimer’s disease. Cell Metab., 31, 503–517.
59) Ni, X. & Mori, H. (2022) Complex processes underlying the dynamic changes of D-serine levels in AD brains. Curr. Alzheimer Res., 19, 485–493.
60) Balu, D.T., Pantazopoulos, H., Huang, C.C.Y., Muszynski, K., Harvey, T.L., Uno, Y., Rorabaugh, J.M., Galloway, C.R., Botz-Zapp, C., Berretta, S., et al. (2019) Neurotoxic astrocytes express the D-serine synthesizing enzyme, serine racemase, in Alzheimer’s disease. Neurobiol. Dis., 130, 104511.
61) Ozaki, H., Inoue, R., Matsushima, T., Sasahara, M., Hayashi, A., & Mori, H. (2018) Serine racemase deletion attenuates neurodegeneration and microvascular damage in diabetic retinopathy. PLoS One, 13, e0190864.
62) Jiang, H., Fang, J., Wu, B., Yin, G., Sun, L., Qu, J., Barger, S.W., & Wu, S. (2011) Overexpression of serine racemase in retina and overproduction of D-serine in eyes of streptozotocin-induced diabetic retinopathy. J. Neuroinflammation, 8, 119.
63) Harai, T., Inoue, R., Fujita, Y., Tanaka, A., Horio, M., Hashimoto, K., Hongou, K., Miyawaki, T., & Mori, H. (2012) Decreased susceptibility to seizures induced by pentylenetetrazole in serine racemase knockout mice. Epilepsy Res., 102, 180–187.
64) Perez, E.J., Tapanes, S.A., Loris, Z.B., Balu, D.T., Sick, T.J., Coyle, J.T., & Liebl, D.J. (2017) Enhanced astrocytic D-serine underlies synaptic damage after traumatic brain injury. J. Clin. Invest., 127, 3114–3125.
65) Tapanes, S.A., Arizanovska, D., Díaz, M.M., Folorunso, O.O., Harvey, T., Brown, S.E., Radzishevsky, I., Close, L.N., Jagid, J.R., Graciolli Cordeiro, J., et al. (2022) Inhibition of glial D-serine release rescues synaptic damage after brain injury. Glia, 70, 1133–1152.
66) Liraz-Zaltsman, S., Slusher, B., Atrakchi-Baranes, D., Rosenblatt, K., Friedman Levi, Y., Kesner, E., Silva, A.J., Biegon, A., & Shohami, E. (2018) Enhancement of brain D-serine mediates recovery of cognitive function after traumatic brain injury. J. Neurotrauma, 35, 1667–1680.
67) Biegon, A., Liraz-Zaltsman, S., & Shohami, E. (2018) Stimulation of N-methyl-D-aspartate receptors by exogenous and endogenous ligands improves outcome of brain injury. Curr. Opin. Neurol., 31, 687–692.
68) Tabata-Imai, A., Inoue, R., & Mori, H. (2014) Increased sensitivity to inflammatory pain induced by subcutaneous formalin injection in serine racemase knock-out mice. PLoS One, 9, e105282.
69) Balan, L., Foltyn, V.N., Zehl, M., Dumin, E., Dikopoltsev, E., Knoh, D., Ohno, Y., Kihara, A., Jensen, O.N., Radzishevsky, I.S., et al. (2009) Feedback inactivation of D-serine synthesis by NMDA receptor-elicited translocation of serine racemase to the membrane. Proc. Natl. Acad. Sci. USA, 106, 7589–7594.
70) Tsuzuki, H., Maekawa, M., Konno, R., & Hori, Y. (2012) Functional roles of endogenous D-serine in pain-induced ultrasonic vocalization. Neuroreport, 23, 937–941.
71) Horio, M., Kohno, M., Fujita, Y., Ishima, T., Inoue, R., Mori, H., & Hashimoto, K. (2011) Levels of D-serine in the brain and peripheral organs of serine racemase (Srr) knock-out mice. Neurochem. Int., 59, 853–859.
72) Ma, M.C., Huang, H.S., Chen, Y.S., & Lee, S.H. (2008) Mechanosensitive N-methyl-D-aspartate receptors contribute to sensory activation in the rat renal pelvis. Hypertension, 52, 938–944.
73) Sasabe, J., Suzuki, M., Miyoshi, Y., Tojo, Y., Okamura, C., Ito, S., Konno, R., Mita, M., Hamase, K., & Aiso, S. (2014) Ischemic acute kidney injury perturbs homeostasis of serine enantiomers in the body fluid in mice: Early detection of renal dysfunction using the ratio of serine enantiomers. PLoS One, 9, e86504.
74) Ganote, C.E., Peterson, D.R., & Carone, F.A. (1974) The nature of D-serine-induced nephrotoxicity. Am. J. Pathol., 77, 269–282.
75) Kimura, T., Hamase, K., Miyoshi, Y., Yamamoto, R., Yasuda, K., Mita, M., Rakugi, H., Hayashi, T., & Isaka, Y. (2016) Chiral amino acid metabolomics for novel biomarker screening in the prognosis of chronic kidney disease. Sci. Rep., 6, 26137.
76) Nakade, Y., Iwata, Y., Furuichi, K., Mita, M., Hamase, K., Konno, R., Miyake, T., Sakai, N., Kitajima, S., Toyama, T., et al. (2018) Gut microbiota-derived D-serine protects against acute kidney injury. JCI Insight, 3, e97957.
77) Inoue, R., Yoshihisa, Y., Tojo, Y., Okamura, C., Yoshida, Y., Kishimoto, J., Luan, X., Watanabe, M., Mizuguchi, M., Nabeshima, Y., et al. (2014) Localization of serine racemase and its role in the skin. J. Invest. Dermatol., 134, 1618–1626.
78) Yoshihisa, Y., Rehman, M.U., Nakagawa, M., Matsukuma, S., Makino, T., Mori, H., & Shimizu, T. (2018) Inflammatory cytokine-mediated induction of serine racemase in atopic dermatitis. J. Cell. Mol. Med., 22, 3133–3138.
79) Montesinos Guevara, C. & Mani, A.R. (2016) The role of D-serine in peripheral tissues. Eur. J. Pharmacol., 780, 216–223.
80) Uda, K., Edashige, Y., Nishimura, R., Shikano, Y., Matsui, T., Radkov, A.D., & Moe, L.A. (2020) Distribution and evolution of the serine/aspartate racemase family in plants. Phytochemistry, 169, 112164.
81) Saitoh, Y., Katane, M., Miyamoto, T., Sekine, M., Sakai-Kato, K., & Homma, H. (2020) D-Serine and D-alanine regulate adaptive foraging behavior in Caenorhabditis elegans via the NMDA receptor. J. Neurosci., 40, 7531–7544.
82) Dai, X., Zhou, E., Yang, W., Zhang, X., Zhang, W., & Rao, Y. (2019) D-Serine made by serine racemase in Drosophila intestine plays a physiological role in sleep. Nat. Commun., 10, 1986.
83) Krasovec, G., Hozumi, A., Yoshida, T., Obita, T., Hamada, M., Shiraishi, A., Satake, H., Horie, T., Mori, H., & Sasakura, Y. (2022) D-Serine controls epidermal vesicle release via NMDA receptor, allowing tissue migration during the metamorphosis of the chordate Ciona. Sci. Adv., 8, eabn3264.
84) Horio, M., Ishima, T., Fujita, Y., Inoue, R., Mori, H., & Hashimoto, K. (2013) Decreased levels of free D-aspartic acid in the forebrain of serine racemase (Srr) knock-out mice. Neurochem. Int., 62, 843–847.
85) Semenza, E.R., Harraz, M.M., Abramson, E., Malla, A.P., Vasavda, C., Gadalla, M.M., Kornberg, M.D., Snyder, S.H., & Roychaudhuri, R. (2021) D-cysteine is an endogenous regulator of neural progenitor cell dynamics in the mammalian brain. Proc. Natl. Acad. Sci. USA, 118, e2110610118.
著者紹介Author Profile
 森 寿(もり ひさし)
森 寿(もり ひさし)富山大学学術研究部医学系医学部分子神経科学講座 教授.医学博士.
略歴1985年広島大学理学部生物学科卒業,91年大阪大学大学院医学研究科博士課程修了,同年新潟大学脳研究所神経薬理学部門助手,94年東京大学医学部薬理学講座助手,99年同講師,2003年富山医科薬科大学医学部生化学第2講座教授,05年より大学統合により富山大学医学部分子神経科学講座教授.
研究テーマと抱負D-アミノ酸の生体機能と代謝機構の解析.
趣味アウトドア(海釣りから山登りまで),犬吸い.