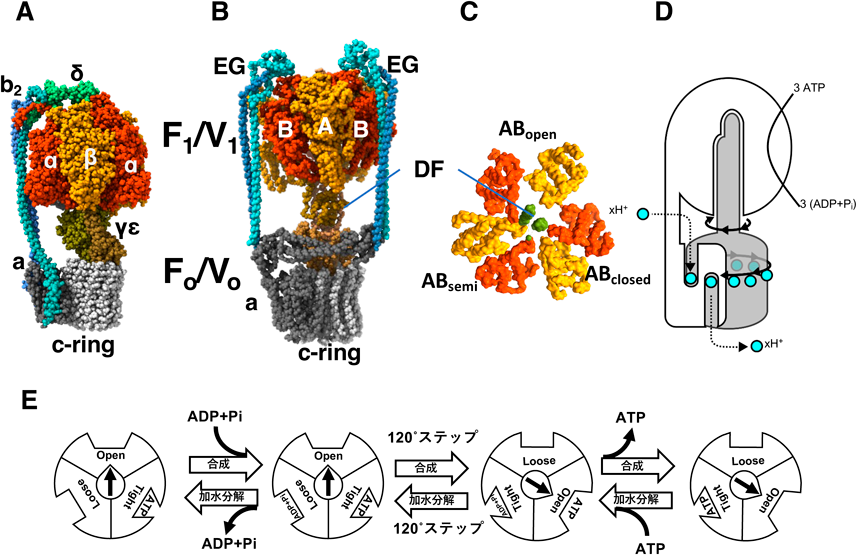
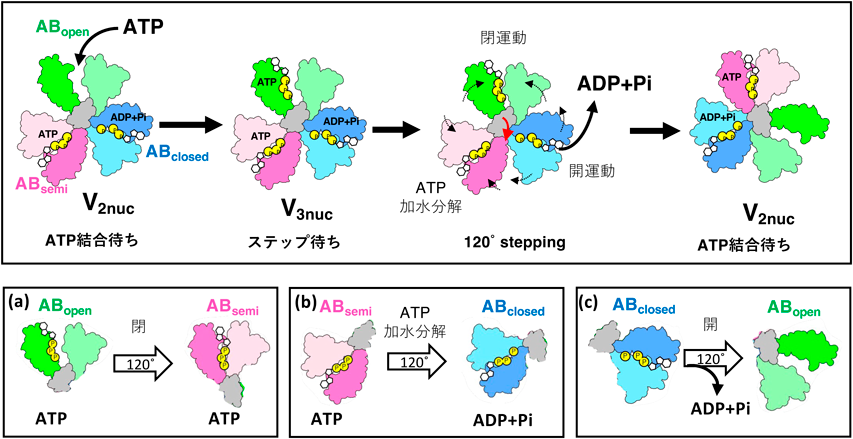
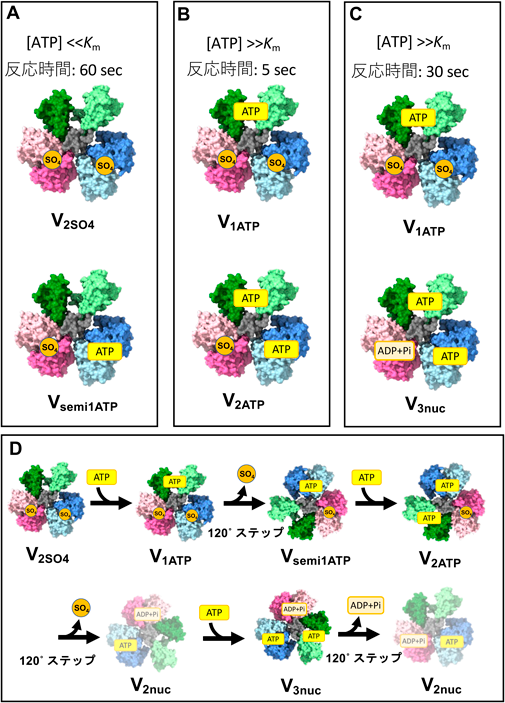
クライオ電子顕微鏡による回転型ATPaseの構造生化学Structural biochemical analysis of rotary ATPases by Cryo electron microscopy
京都産業大学 生命科学部Kyoto Sangyo University, Faculty of Biosciences ◇ 〒603–8555 京都市北区上賀茂本山 生命科学部16号館 ◇ Kamigamo Mitoyama, Kita-ku, Kyoto 603–8555, Japan
発行日:2023年8月25日Published: August 25, 2023