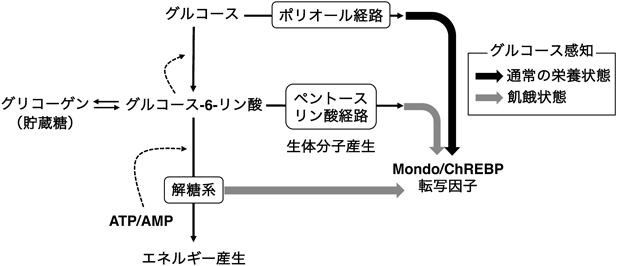
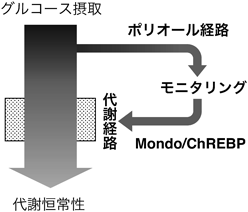
グルコース代謝経路である「ポリオール経路」による進化的に保存されたグルコース感知機構Evolutionarily conserved role of the polyol pathway in sensing glucose uptake
久留米大学分子生命科学研究所Institute of Life Science, Kurume University ◇ 福岡県久留米市旭町67番地 ◇ 67 Asahi-machi, Kurume, Fukuoka 830–0011, Japan
発行日:2023年8月25日Published: August 25, 2023