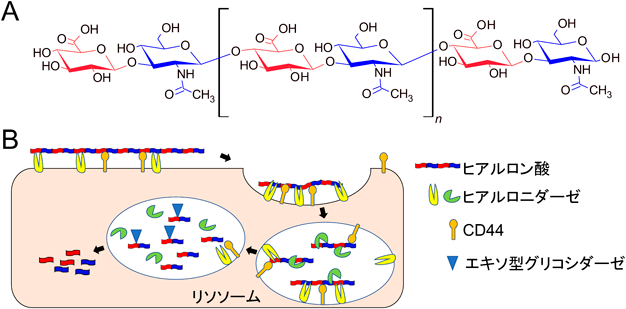
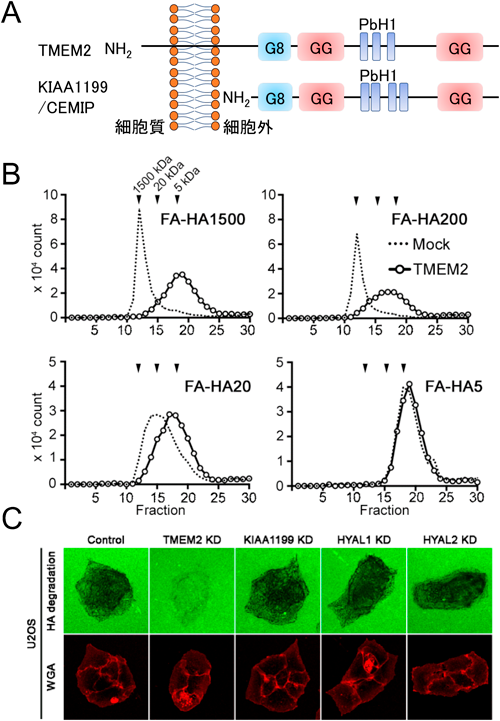
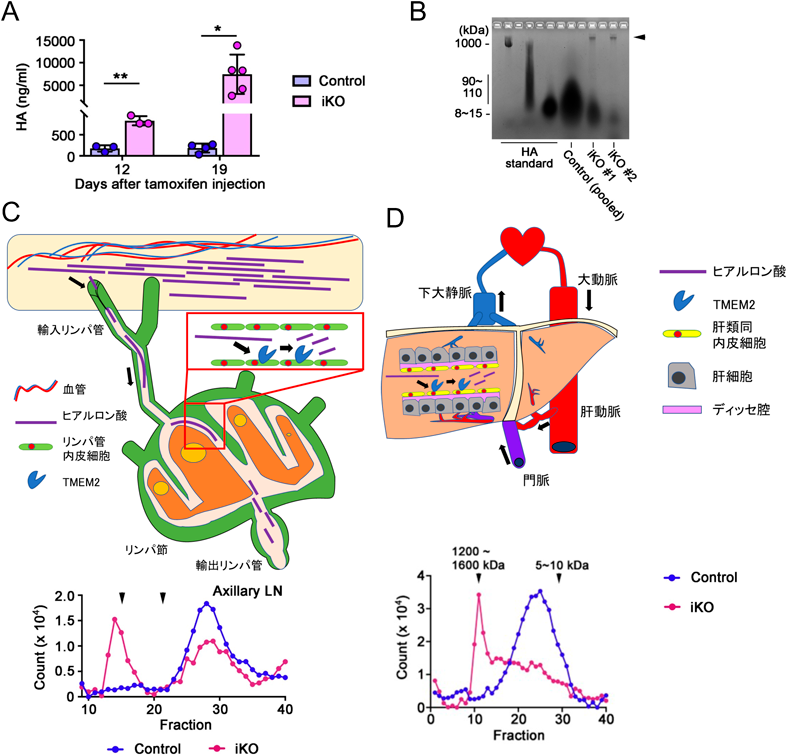
TMEM2による個体レベルでのヒアルロン酸分解機構TMEM2 is important for systemic hyaluronan metabolism at the individual level
岐阜大学医学部附属病院・泌尿器科Department of Urology, Gifu University Hospital ◇ 〒501–1194 岐阜県岐阜市柳戸1–1 ◇ Yanagido 1–1, Gifu, Gifu 501–1194, Japan
発行日:2023年8月25日Published: August 25, 2023