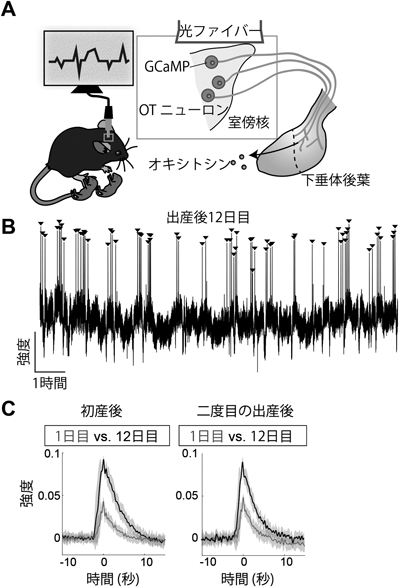
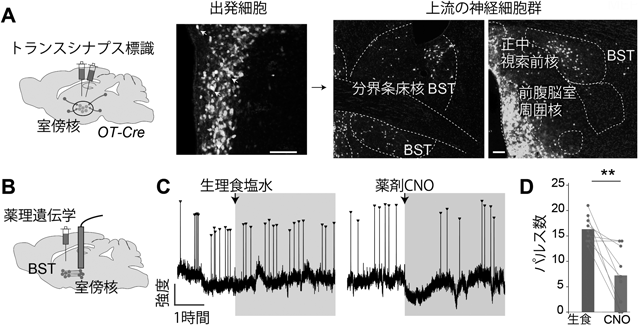
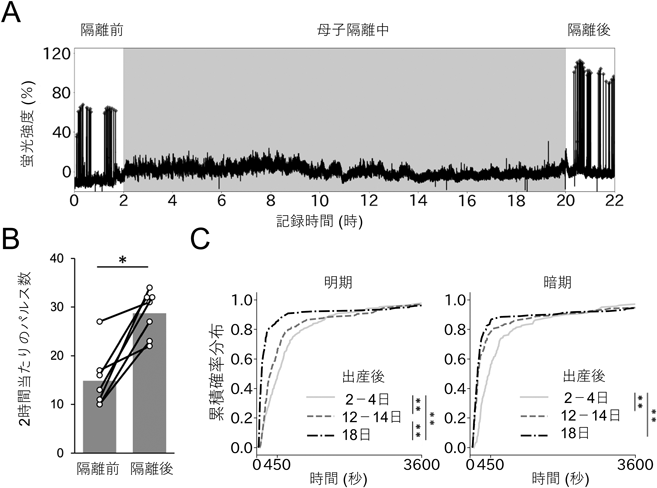
授乳期マウスのオキシトシン分泌動態を可視化するRecording of pulsatile activities of oxytocin neurons in lactating mice
矢口 花紗音,幸長 弘子,宮道 和成 Kasane Yaguchi, Hiroko Yukinaga, Kazunari Miyamichi
Kasane Yaguchi, Hiroko Yukinaga, Kazunari Miyamichi
Kasane Yaguchi, Hiroko Yukinaga, Kazunari Miyamichi理化学研究所 生命機能科学研究センターRIKEN Biosystems Dynamics Research ◇ 〒650–0047 兵庫県神戸市中央区港島中町2–2–3 ◇ 2–2–3 Minatojima-mimamicho, Chuo-ku, Kobe, Hyogo 650–0047, Japan
発行日:2023年10月25日Published: October 25, 2023