CRISPR-CasとOMEGAシステムの分子基盤Molecular basis of CRISPR-Cas and OMEGA systems
ブロード研究所Broad Institute ◇ マサチューセッツ州ケンブリッジ市エイムス街75番地 ◇ 75 Ames street, Cambridge, MA, USA
発行日:2023年10月25日Published: October 25, 2023
© 2023 公益社団法人日本生化学会© 2023 The Japanese Biochemical Society
生物のゲノムにはさまざまな機能を持ったシステムがコードされており,人類に有用な基盤技術として利用されることがある.本稿では,生命科学を中心としたさまざまな分野に影響を与えるCRISPR-Casと呼ばれるシステム,および,CRISPR-Casとともに注目を集めるOMEGAと呼ばれるシステムについて,それぞれの分子基盤を解説する.
CRISPR(clustered regularly interspaced short palindromic repeats)-Cas(CRISPR associated)システムは原核生物に備わる生体防御機構である1)(図1A).宿主がウイルスやプラスミドに侵入されたとき,侵入者の配列情報はCRISPR-Casシステムに記録される.宿主は記録された配列情報とCasタンパク質を用いて,侵入者を無力化する.このように,宿主はCRISPR-Casシステムを用いて,侵入者の配列情報から免疫を獲得する.
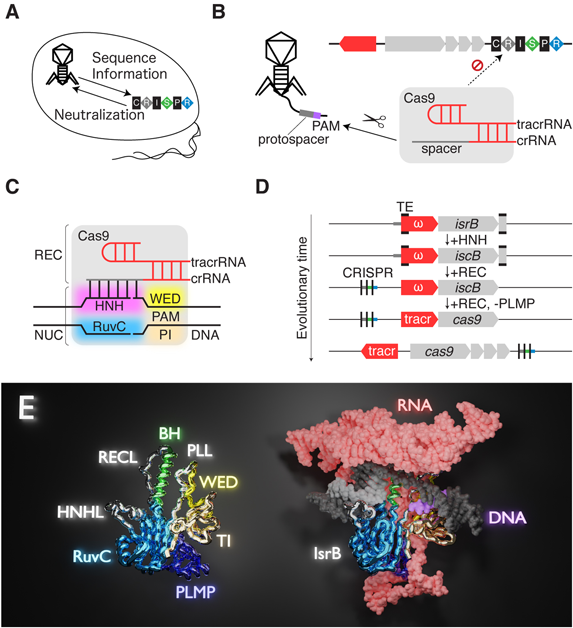
(A)生体防御機構CRISPR-Casシステム.(B)II型のCRISPR-Casシステム.(C)Cas9の作動機構.(D)Cas9と関連する遺伝子座の変遷.TE:トランスポゾン末端領域.ω:ωRNAをコードする領域.(E)IsrB–ωRNA–標的DNA複合体の構造.ωRNAのガイド配列,標的DNAのTAM配列を灰色,紫色で示した.
後述するOMEGAシステムと関連が深いII型のCRISPR-Casシステムでは,Cas9タンパク質が侵入者のDNA(以下,外来DNAと呼ぶ)を切断する1)(図1B).Cas9は,外来DNAの配列情報を含むcrRNA(CRISPR RNA)と機能的な足場を形成するtracrRNA(trans-activating crRNA)と協働して,外来DNAを標的とする.このとき,標的配列(protospacer)がcrRNAのガイド配列(spacer)と相補性を持つことに加えて,PAM(protospacer adjacent motif)と呼ばれる塩基モチーフが標的配列の近傍にあることが必要である.CRISPR-Casシステムに記録する外来DNAを選ぶときはPAMを近傍に持つようにしている一方で,記録するときにはPAMはトリミングされる2, 3).したがって,Cas9は外来DNAを記録した宿主のDNA(PAMを持たない)は認識せず,外来DNA(PAMを持つ)だけを標的とし,切断する.このように,Cas9はII型のCRISPR-Casシステムで中心的役割を果たす.
Cas9の作動機構が生化学解析,構造解析,動態解析より明らかになった4)(図1C).Cas9は標的DNAを認識するRECローブと標的DNAを切断するNUCローブに分かれ,RECローブはαヘリックスに富んだCas9独自のフォールディングを形成し,NUCローブは二つのヌクレアーゼドメイン(RuvC, HNHドメイン)と二つの新規ドメイン(WED, PIドメイン)から構成される.Cas9がWEDとPIドメインによって形成される溝で標的DNAのPAM配列を認識した後,Cas9と協働するcrRNAが標的DNAと塩基対を形成する.crRNAと標的DNAで形成されるRNA-DNAヘテロ二本鎖がRECローブによって認識されることで,NUCローブの二つのヌクレアーゼドメインが活性化されて標的DNAの二本鎖切断が行われる.
Cas9, crRNA, tracrRNAのエンジニアリングによってCas9は生命科学と医療の分野で応用されるようになった1).crRNAとtracrRNAからなるガイドRNAとCas9を導入することによってヒト細胞やモデル生物のゲノムを編集することができる.Cas9がゲノムの標的部位を二本鎖切断した後,DNA修復によってランダムに塩基を挿入,欠損させたり,特定の塩基配列を挿入したりすることができる.Cas9の二つのヌクレアーゼドメインの両方を不活性化したdCas9に各種エフェクタータンパク質を融合,リクルートすることによって,ゲノムの標的部位を転写制御,エピジェネティック修飾することができる.Cas9の片方のヌクレアーゼドメインを不活性化したCas9ニッカーゼに脱アミノ化酵素や逆転写酵素を融合させることによって,ゲノムの標的部位の塩基編集やプライム編集を行える5).Cas9を用いたゲノム編集により,疾患モデル細胞,動物が簡便に作製できるようになった他,病原性変異修復,有用変異導入,免疫細胞改変による治療が可能となった.実例として,鎌状赤血球症患者に対し,BCL11A遺伝子のエンハンサー領域をCas9でゲノム編集した造血幹細胞を移植して有効な結果を得ている6).
近年,ゲノミクスの分野において,細菌のゲノムが年間数千件,メタゲノムではさらに多くのデータが公共レポジトリに登録され続けており,技術革新のための生体高分子が探索されている.Cas9の特徴であるブリッジヘリックス(REC, NUCローブを連結するαヘリックス,BH),および,二つのヌクレアーゼドメイン(RuvCとHNH)を持つIscB(insertion sequences Cas9-like ORF B)と呼ばれるタンパク質がIS200/605トランスポゾンにコードされていることが発見され,Cas9の祖先にあたると推定された7).このトランスポゾンからRuvCドメインとBHを持つIsrB(insertion sequences RuvC-like ORF B)も発見された.
当研究室はIscB, IsrBとノンコーディングRNAの関連および機能を解明した8)(図1D).crRNAがコードされているCRISPRアレイを近傍に持つiscB遺伝子を発見し,この遺伝子産物をCasIscB(CRISPR-associated IscB)と命名した.CasIscBはCRISPRアレイとiscB遺伝子の中間領域より発現するノンコーディングRNAとcrRNAに関わり,Cas9のように,crRNAのガイド配列を利用して標的DNAを切断する.CRISPRアレイを持たないiscB遺伝子座ではIscBと関わるRNAがトランスポゾンの末端領域からiscB遺伝子にかけてコードされ,iscB遺伝子を持つ微生物で発現している.これをωRNAと命名した.CasIscBのRNAとωRNAは同様の二次構造を持ち,特にガイド配列の近傍の二本鎖領域が保存されている9).当研究室はωRNAにガイド配列を追加することで,IscBが任意のDNAを切断することができることを示した8).同様に,IsrBはガイド配列の追加されたωRNAを利用して標的DNAを一本鎖切断し,ニックを導入することができる.IscB, IsrBはCas9にはないPLMPドメインを持ち,TAM(target adjacent motif)と呼ばれる塩基モチーフを含む標的DNAを,それぞれ二本鎖切断,一本鎖切断することができる.このような特徴を持ち,ωRNAとともにトランスポゾンにコードされたRNA依存性エンドヌクレアーゼをOMEGA(obligate mobile element guided activity)システムと命名した.
CRISPR-CasとOMEGAシステムの進化的関連を調べるため,IscB, IsrB, Cas9を系統解析したところ,三つのヌクレアーゼは共通の進化系譜より発生したことがわかった8).IsrBは系統樹における発生初期の位置にあり,ほとんどすべてのIsrBがωRNAを持っていたことから,OMEGAシステムは,RuvCエンドヌクレアーゼが挿入配列およびωRNAを得て進化したRNA依存性エンドヌクレアーゼIsrBに始まり,HNHドメインの獲得によって進化したIscBへ変遷したことが示唆された.IscBの進化の中でCasIscBはまれに現れ,このときCasIscBへのRECローブ様ドメイン挿入とCRISPRアレイとの相関が同時に起こっている.PLMPドメインを欠いたCasIscBからCas9ファミリーが発生した.発生初期のCas9のtracrRNAが有意にCasIscBのωRNAと類似していたことから,tracrRNAはωRNAから進化したことが示唆された.このように,OMEGAからCRISPR-Casシステムにかけて,エフェクターのヌクレアーゼとそれに関わるRNAが共進化していることが明らかになった.
IsrBは,Cas9と異なるドメイン,RNA構造を持つ一方で,Cas9と同様のRNA依存的なDNA標的機構を成立させており,その分子機構は不明であった.そこで著者らは,IsrB, ωRNA,標的DNAからなる複合体の立体構造を決定した10)(図1E).立体構造と変異体解析の結果から,IsrBの機能性ドメイン,モチーフを決定した.RuvCドメインは,三つの触媒モチーフ(RuvC I–III)の間に,三つの配列(BH, A, B)を持つ.配列Aは,BHとRuvC IIの間をわずか17残基でショートカットするリンカーであり,Cas9では350から800残基ほどのRECローブに置き換えられるため,RECLと命名した.RuvC IIとIIIの間にある配列Bは,ループとαヘリックスからなる単純なリンカーで,Cas9ではHNHドメインに置き換えられるため,HNHLと命名した.C末端の三つの領域はTAMを含むDNA二重鎖に結合しており,Cas9と類似の機能を持っていたため,PLL(phosphate lock loop),WEDドメイン,TI(TAM-interacting)ドメインと命名した.PLMPドメインは,βシートとαヘリックスからなり,翻訳開始因子IF3のN末端ドメインと似た構造を持つ11).PLMPドメインは,RuvCドメインとTIドメインの両方と広範囲に相互作用しており,その機能を支えることが示唆された.
ωRNAは,標的DNAと対合するガイド部位とスカフォールド部位から構成されている.ωRNAスカフォールドでは,10個のRNAヘリックスがシュードノットやスタッキング相互作用を介して球状構造をとっていた.ωRNAが自律的に球状構造を形成し,足場として機能すれば,エフェクタータンパク質はRNAの折りたたみや機能をサポートする補助モチーフやドメインを必要としないため,OMEGAシステムにとって有益であると考えられる.さらに,もし球状構造がRNA分解に対する抵抗力を持つなら,ωRNAがエフェクタータンパク質とトランスで機能することを促進する可能性もある.この可能性は,異なる遺伝子座にコードされたIscBと機能することができる自律的なωRNAの発見によって支持されている8).ωRNAの5′末端ステム領域には,シュードノットを形成するヘアピンが挿入されており,球状構造形成に寄与していた.CRISPR-Casシステムへの進化の過程で,5′末端ステム領域がCRISPRアレイの配列に適合することで,機能的なCas9-crRNA-tracrRNA複合体の形成が可能になったと考えられる.ωRNAの3′末端はIsrBのN末端に向かってのびており,二つのヘアピンがPLMPドメインと相互作用していた.このヘアピンにはIsrBコード領域上流のShine-Dalgarno配列が含まれていることから,IsrBとωRNAの相互作用がIsrB発現制御に寄与する可能性が示された.
IsrBの構造から,RNA依存的なDNA標的機構の起源が明らかになった.さらに,IscB, Cas9との構造比較から,OMEGAからCRISPR-Casシステムにかけてどのようにこの機構が変遷したか明らかになった12, 13)(図2).まず,RuvCヌクレアーゼにさまざまなモチーフ,ドメインが挿入されたIsrBが,ωRNAと協働することによって,DNAを標的とすることができるようになった.WED, TIドメインがDNAの塩基モチーフを認識し,PLLやBHと協働することによってDNAをほどくことができる.このとき,ωRNAスカフォールドが直接DNAを認識することによって標的DNAが選択される.IscBではωRNAスカフォールドが縮退し,HNHドメインを獲得することによって,よりタンパク質に依存したDNA標的機構へ移行する.Cas9ではRNAの構成が変わり,RECローブを獲得することによってタンパク質によるDNA標的機構が確立する.以上,立体構造からCRISPR-Casの進化について理解が深まった.OMEGAシステムはかさ高いRNAを利用することによって,エフェクターヌクレアーゼの機能をRNAに移行し,ヌクレアーゼの小型化を可能にしている.今後はIsrB, IscBの構造情報を利用して,塩基編集,プライム編集のためのエフェクタータンパク質を開発し,生命科学や医療に有用な基盤技術を発展させることが期待される14).
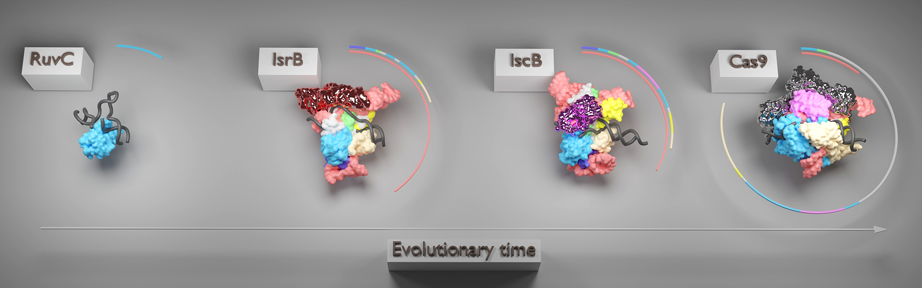
PDB番号6S16(RuvC), 8CSZ(IscB), 7S4X(Cas9)を使用.円グラフでタンパク質,RNAの分子量を示した.分子進化で獲得され,ヌクレアーゼの機能を改変,向上させた領域を光沢で示した.
1) Zhang, F. (2019) Development of CRISPR-Cas systems for genome editing and beyond. Q. Rev. Biophys., 52, e6–e6.
2) Heler, R., Samai, P., Modell, J.W., Weiner, C., Goldberg, G.W., Bikard, D., & Marraffini, L.A. (2015) Cas9 specifies functional viral targets during CRISPR-Cas adaptation. Nature, 519, 199–202.
3) Wang, J.Y., Tuck, O.T., Skopintsev, P., Soczek, K.M., Li, G., Al-Shayeb, B., Zhou, J., & Doudna, J.A. (2023) Genome expansion by a CRISPR trimmer-integrase. Nature, 618, 855–861.
4) Wang, J.Y., Pausch, P., & Doudna, J.A. (2022) Structural biology of CRISPR-Cas immunity and genome editing enzymes. Nat. Rev. Microbiol., 20, 641–656.
5) Anzalone, A.V., Randolph, P.B., Davis, J.R., Sousa, A.A., Koblan, L.W., Levy, J.M., Chen, P.J., Wilson, C., Newby, G.A., Raguram, A., et al. (2019) Search-and-replace genome editing without double-strand breaks or donor DNA. Nature, 576, 149–157.
6) Mehta, J. (2021) CRISPR-Cas9 gene editing for sickle cell disease and β-thalassemia. N. Engl. J. Med., 384, e91.
7) Kapitonov, V.V., Makarova, K.S., & Koonin, E.V. (2015) ISC, a novel group of bacterial and archaeal DNA transposons that encode Cas9 homologs. J. Bacteriol., 198, 797–807.
8) Altae-Tran, H., Kannan, S., Demircioglu, F.E., Oshiro, R., Nety, S.P., McKay, L.J., Dlakić, M., Inskeep, W.P., Makarova, K.S., Macrae, R.K., et al. (2021) The widespread IS200/IS605 transposon family encodes diverse programmable RNA-guided endonucleases. Science, 374, 57–65.
9) Weinberg, Z., Perreault, J., Meyer, M.M., & Breaker, R.R. (2009) Exceptional structured noncoding RNAs revealed by bacterial metagenome analysis. Nature, 462, 656–659.
10) Hirano, S., Kappel, K., Altae-Tran, H., Faure, G., Wilkinson, M.E., Kannan, S., Demircioglu, F.E., Yan, R., Shiozaki, M., Yu, Z., et al. (2022) Structure of the OMEGA nickase IsrB in complex with ωRNA and target DNA. Nature, 610, 575–581.
11) Biou, V., Shu, F., & Ramakrishnan, V. (1995) X-ray crystallography shows that translational initiation factor IF3 consists of two compact alpha/beta domains linked by an alpha-helix. EMBO J., 14, 4056–4064.
12) Schuler, G., Hu, C., & Ke, A. (2022) Structural basis for RNA-guided DNA cleavage by IscB-ωRNA and mechanistic comparison with Cas9. Science, 376, 1476–1481.
13) Bravo, J.P.K., Liu, M.S., Hibshman, G.N., Dangerfield, T.L., Jung, K., McCool, R.S., Johnson, K.A., & Taylor, D.W. (2022) Structural basis for mismatch surveillance by CRISPR-Cas9. Nature, 603, 343–347.
14) Aliaga Goltsman, D.S., Alexander, L.M., Lin, J.L., Fregoso Ocampo, R., Freeman, B., Lamothe, R.C., Perez Rivas, A., Temoche-Diaz, M.M., Chadha, S., Nordenfelt, N., et al. (2022) Compact Cas9d and HEARO enzymes for genome editing discovered from uncultivated microbes. Nat. Commun., 13, 7602.

ブロード研究所 博士研究員.博士(理学).
2020年3月東京大学大学院理学系研究科生物科学専攻にて博士号取得.同年7月より現職.
研究テーマCRISPR-Cas9の構造と機能.

This page was created on 2023-09-08T15:30:11.876+09:00
This page was last modified on 2023-10-11T08:29:00.000+09:00
このサイトは(株)国際文献社によって運用されています。