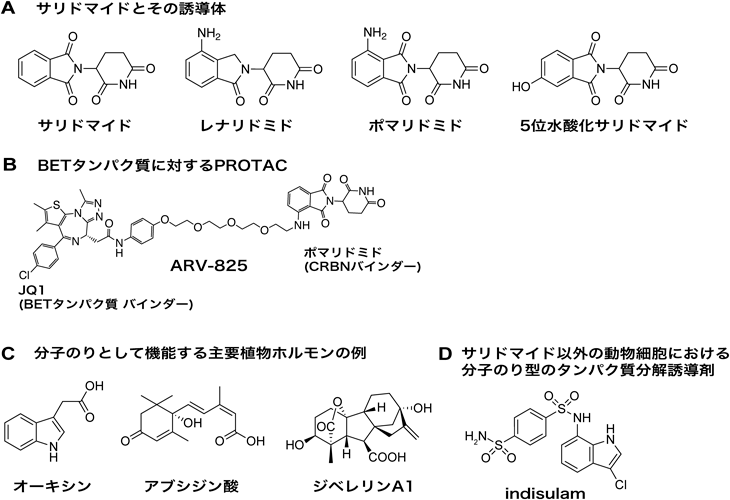
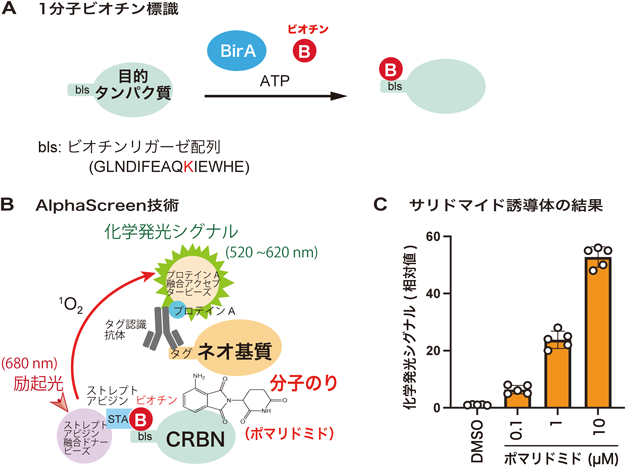
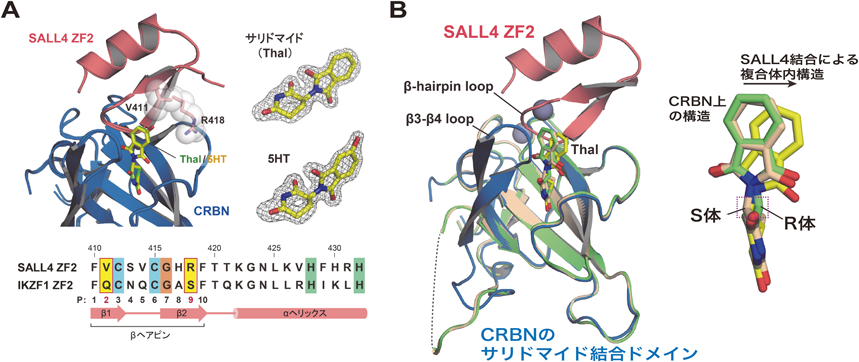
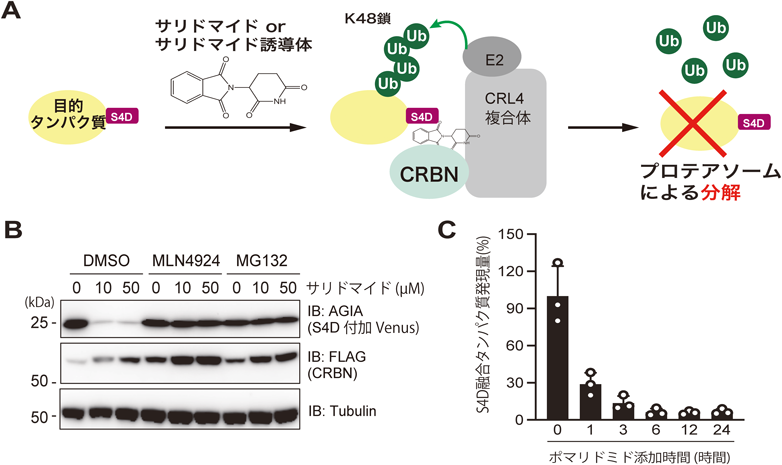
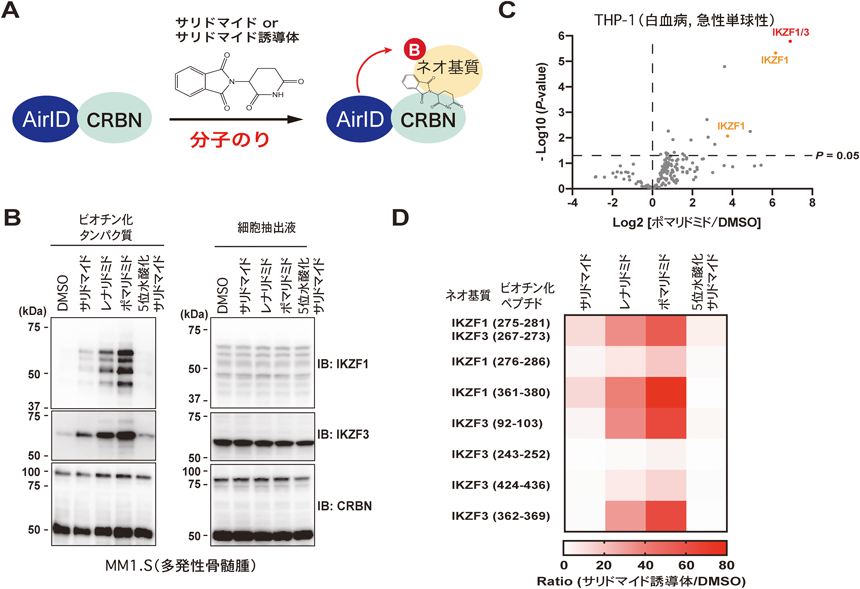
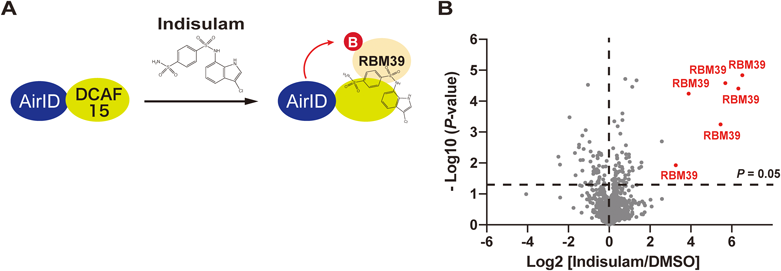
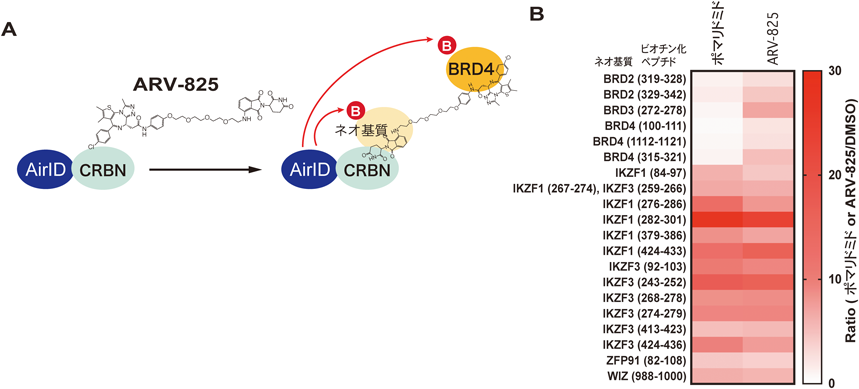
薬剤依存的にプロテアソームにより分解されるタンパク質のための新技術New technology for analysis of drug-dependently degraded proteins by proteasome
愛媛大学プロテオサイエンスセンターProteo-Science Center, Ehime University ◇ 〒790–8577 愛媛県松山市文京町3 ◇ 3 Bunkyo-cho, Matsuyama, Ehime 790–8577, Japan
発行日:2023年12月25日Published: December 25, 2023