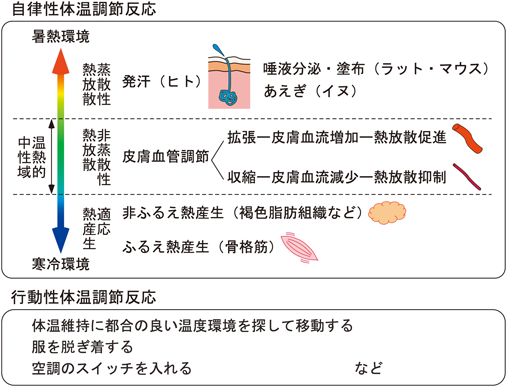
哺乳類の体温調節反応は自律性体温調節反応と行動性体温調節反応に大別される(図1).自律性体温調節反応は意志や意識の有無とは関係なく起こる自律的な生理反応であり,代表的なものとして,ふるえ熱産生,非ふるえ(代謝)熱産生,皮膚血管調節があげられる.ヒトでは発汗も重要な自律性体温調節反応である.このうち,非ふるえ熱産生が起こる主要な器官として褐色脂肪組織が知られる.褐色脂肪組織は交感神経の支配を受け,神経終末から放出されるノルアドレナリンや副腎髄質由来のアドレナリンがβ3アドレナリン受容体を介して褐色脂肪細胞に作用する.すると,脂肪細胞内でミトコンドリアの脱共役タンパク質(uncoupling protein 1)が活性化され,それが電子伝達系とアデノシン5′-三リン酸(ATP)合成機構とを脱共役させることによって,ATP合成に向かうエネルギーが熱エネルギーとして放出されることで熱産生が起こる3).
皮膚血管も主に交感神経の支配を受け,交感神経入力によって血管平滑筋が収縮すると皮膚血流が低下し,体表面から環境中への体熱の放散が抑制される.一方,交感神経入力が低下すると血管平滑筋が弛緩して皮膚血流が増加するため,体熱の放散が促進される(図1).ふるえ熱産生は骨格筋が高頻度に収縮と弛緩を繰り返すことによって大量の熱を産生させる反応であり,体性運動神経を通じて起こる4).発汗は,アセチルコリンを放出する特殊な交感神経によって支配されるエクリン腺から汗が皮膚表面に分泌される現象であり,汗の蒸発に伴う気化熱として体熱が奪われることにより熱の放散を促進する.実験動物として用いられるラットやマウスなどは体温調節の発汗機能を持たないため,発汗を制御する中枢神経メカニズムの研究はあまり進んでいない.
皮膚血管の制御などによる非蒸散性熱放散(汗などの水分の蒸発を伴わない熱放散)の量を調節するだけで深部体温を維持することができる環境温度の範囲を温熱的中性域と呼び,その温度域の範囲内で環境温度が低下すると皮膚血管が収縮して熱放散を抑制する.温熱的中性域よりもさらに環境温度が低下すると適応熱産生と呼ばれる積極的な熱産生が起こるようになり,まず,褐色脂肪熱産生が惹起される(図1).それでも体温低下を防ぐことができない程度に環境温度が低下すると,ふるえ熱産生が起こる5).一方,温熱的中性域の範囲内で環境温度が上昇すると皮膚血管が拡張して非蒸散性熱放散を促進するが,温熱的中性域を超えて環境温度が上昇すると,発汗などの蒸散性熱放散反応を起こすことで体温上昇を防ごうとする.ラットやマウスでは唾液の分泌が亢進して体表面に塗布する行動がみられ,イヌなどではあえぎ(パンティング,浅速呼吸)が起こって気道上皮の水分の蒸散を促進する(図1).
こうした自律性体温調節反応には血液循環パラメーターの変化も伴う.たとえば,ラットの皮膚を冷却すると適応熱産生が惹起されるとともに脈拍が上昇する6).また,感染による発熱時には脈拍だけでなく血圧も上昇する7).こうした循環器反応は,熱産生に必要なエネルギー源(糖や脂肪酸など)や酸素の熱産生器官への供給を増やすとともに,産生された熱の全身への分配を促進する効果があると考えられる.また,マウスのような小型動物では,心拍数の上昇に伴って増大する心臓の熱産生が寒冷環境での体温維持や感染時の発熱に寄与する可能性も指摘されている8).
一方,行動性体温調節反応は,体温調節を目的とした意志に基づく行動であり,たとえば,暑い日に木陰を探して涼む,寒いときに上着を羽織る,エアコンや暖房のスイッチを入れるなどの行動があげられる(図1).こうした行動は,自律性体温調節に都合のよい環境に身を置くことで,体温調節に必要なエネルギーや水分といった生体内のリソースの消耗を軽減する効果がある.体温調節行動は本能行動の一つであり,温度感覚によって生まれる快・不快情動がその行動を駆動すると考えられている9).なお,変温動物では行動性体温調節が主体となる(例:カメの甲羅干し).
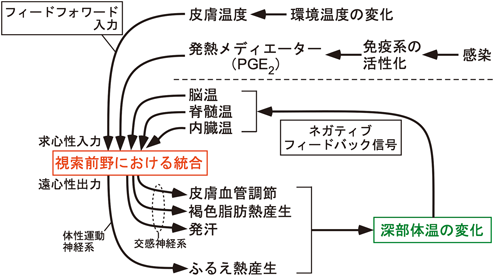
体温の調節には深部体温と環境温度の情報が必要である.視床下部の視索前野にある体温調節中枢にはこの二つの情報がもたらされ,統合される(図2).深部体温の受容器は脳,脊髄,腹腔などに存在するが,視索前野自体の中に存在する温度感受性ニューロンによる脳温の感知が重要な役割を担うと考えられている.深部体温が変動したとき,深部体温を設定範囲へ戻すための体温調節反応が起こるが,これはネガティブフィードバック反応である.しかし,深部体温がいったん変動してから反応を惹起するため,この仕組みだけでは環境温度が変化する中で体温の変動を防ぐことは難しい.そこで生体には,環境温度変化を皮膚で感知し,その情報を神経伝達を介して体温調節中枢へ即座に伝えることで,環境温度変化によって深部体温が変動してしまう前に予防的な体温調節反応を惹起させるフィードフォワード(予測制御)反応の仕組みがある10)(図2).本節では,ネガティブフィードバック反応に必要な情報として深部体温をモニターする温度感受性ニューロンと,フィードフォワード反応に必要な環境温度情報(皮膚温度の感覚情報)を視索前野へ伝える神経路について説明する.
1)温度感受性ニューロン
視索前野には脳組織の温度変化に応じて内因性の発火(シナプス入力を遮断しても起こる自発発火)の頻度を変化させる温度感受性ニューロンが存在し11),その多くは温度上昇によって発火頻度が増加する温ニューロン(warm-sensitive neuron)である12).温ニューロンの発火頻度は皮膚の温度変化によっても変化することから13),温ニューロンが感知する深部体温の情報は,後述する皮膚から視索前野へ伝達される環境温度情報と,温ニューロンの上で統合されると考えられる.温ニューロンが温度を受容するセンサー分子としてはTRPM2やTRPC4が示唆されているが14, 15),いずれも決め手に欠け,温度感受性の分子メカニズムはいまだ不明である.
2)皮膚からの温度感覚神経路
皮膚の一次求心性神経には,温度感受性を持つ陽イオンチャネルとして知られるTRPファミリーが発現しているが16),TRPチャネルの欠損マウスの中で明確な体温調節異常を示すものが報告されていないことから,体温調節に必要な環境温度情報をモニターする皮膚の温度受容器分子についても不明な点が多い.しかし,メントール受容体TRPM8は寒冷防御に必要な冷受容器の有力候補であり,一方,TRPM2やカプサイシン受容体TRPV1は暑熱防御に必要な温覚に関与している可能性が考えられている17).
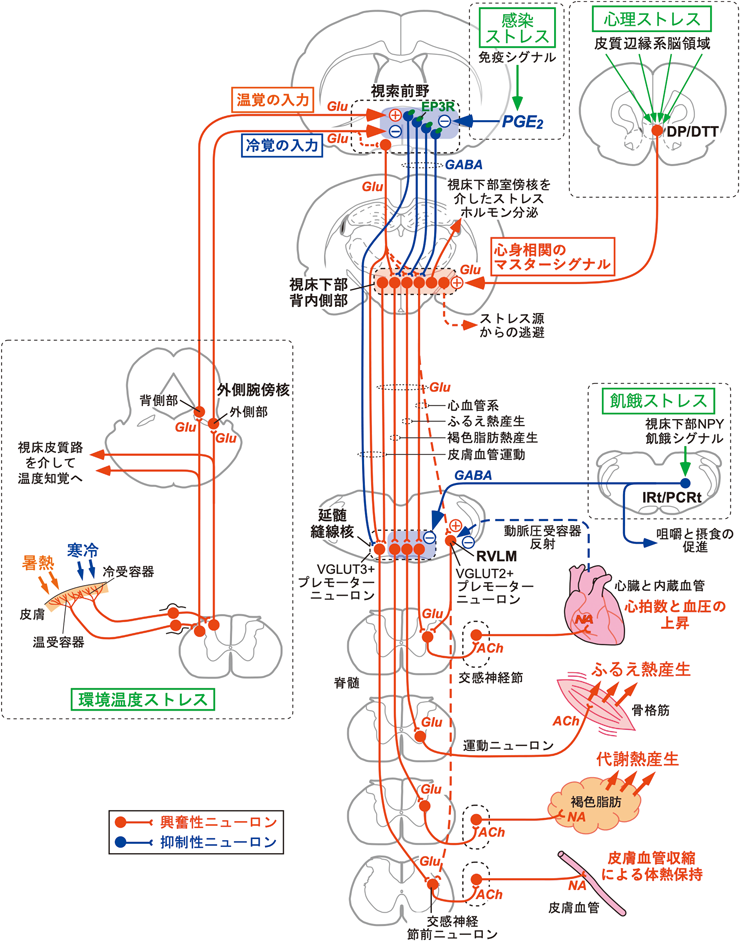
痛みを伴わない温覚と冷覚のシグナルは,異なる一次求心性神経を伝って脊髄あるいは延髄の後角に送られた後,視床と外側腕傍核(lateral parabrachial nucleus)という二つの主要な体性感覚脳領域に中継される18, 19)(図3).視床へ入力された温度感覚シグナルは大脳皮質一次体性感覚野へ送られ,意識に上る皮膚温度の知覚と部位識別のための情報処理に用いられる20).しかし,ラットの視床を破壊し,脊髄視床皮質路を遮断しても自律性ならびに行動性体温調節には影響がないことから21, 22),体温調節に必要な温度感覚メカニズムは,意識の上で温度を知覚するメカニズムとは独立していることがわかる.
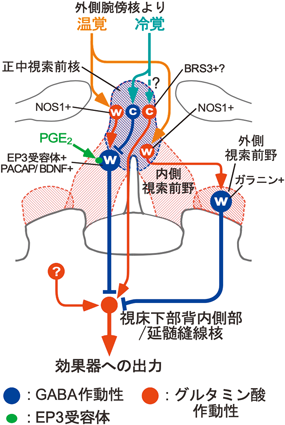
一方,外側腕傍核へ入力される温度感覚情報が体温調節に必要であることが明らかとなった.皮膚由来の冷覚は外側腕傍核の外側部に入力され,温覚は背側部へ入力される21, 23).そして,それぞれの部位から視索前野へ軸索投射するニューロン群を活性化する21, 23)(図3).視索前野へ外側腕傍核外側部から冷覚を伝達するニューロンと背側部から温覚を伝達するニューロンはどちらも転写因子FoxP2を発現するグルタミン酸作動性[小胞型グルタミン酸トランスポーター(VGLUT)2陽性]ニューロンである21, 23–25).外側腕傍核から入力される冷覚と温覚はそれぞれ,視索前野の異なるニューロン群を活性化して異なる体温調節反応を惹起すると考えられる(図4).外側腕傍核外側部から視索前野へ入力される冷覚シグナルは環境温度低下に対する褐色脂肪熱産生や骨格筋のふるえ熱産生の惹起に必要であり21, 26–28),外側腕傍核背側部から視索前野へ入力される温覚シグナルは環境温度上昇に対する皮膚血管拡張(熱放散反応)の惹起に必要である23, 28)(図3).最近では,外側腕傍核背側部から視索前野へ温覚を伝達するニューロン群が,神経ペプチドであるプロダイノルフィン,プロエンケファリン,コレシストキニンを発現することが報告され,これらの遺伝子を発現する外側腕傍核ニューロン群の活動操作実験により,この神経路の暑熱防御への重要性が確認された24, 29–31).
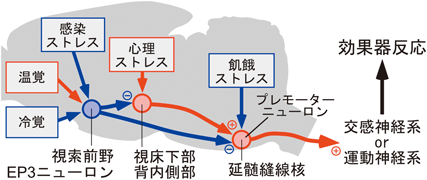
視索前野にある体温調節中枢は,温度感受性ニューロンがモニターする深部体温の情報と,外側腕傍核を介して視索前野へ入力される環境温度情報とを統合し,生体恒常性を維持する上で適切な体温調節を司令すべく末梢効果器へ向けて遠心性シグナルを出力する(図2).視索前野からの遠心路を中継する脳領域として視床下部背内側部(dorsomedial hypothalamus)と吻側延髄縫線核領域(rostral medullary raphe region;以下,延髄縫線核)が知られる(図3,図5).視床下部背内側部から延髄縫線核へはグルタミン酸作動性の興奮性神経伝達が行われ,交感神経出力やふるえ熱産生のための運動神経出力を駆動する27, 33).この興奮性神経路に対し,上位の視索前野から主にGABA作動性の抑制シグナルが恒常的に送られ,その抑制の強さを変えることにより,延髄縫線核から末梢効果器への出力の強さを調節する34).また後述するように,この遠心路には感染ストレス,心理ストレス,飢餓ストレスといった環境ストレスに起因するシグナルが入力し,体温を平熱域から能動的に逸脱させることで環境ストレスから生命を守るためにも機能する(図3,図5).
体温調節の中枢遠心路は,延髄縫線核の交感神経プレモーターニューロン群の同定がきっかけとなって解明が飛躍的に進んだ.本節では,延髄縫線核,視床下部背内側部,そして視索前野のニューロン群の特徴と体温調節司令の伝達メカニズムについて解説する.
1)延髄縫線核の交感神経プレモーターニューロン
主要な体温調節効果器である褐色脂肪組織や皮膚血管は交感神経によって支配される.脊髄からの交感神経司令は胸髄と腰髄の中間質外側核に局在するコリン作動性の交感神経節前ニューロンから出力され,交感神経節のノルアドレナリン作動性の交感神経節後ニューロンを介して効果器へと至る(図3).脊髄の交感神経節前ニューロンに対して脳から興奮性シグナルを伝達するニューロンを交感神経プレモーターニューロンと呼ぶ.
脳内の局所を刺激する生理実験を通じて,褐色脂肪熱産生と皮膚血管収縮を駆動する交感神経プレモーターニューロン群が吻側淡蒼縫線核(rostral medullary raphe nucleus)と大縫線核(raphe magnus nucleus)を中心とする延髄縫線核に分布することが推定されていたが35, 36),2004年にVGLUT3を発現するグルタミン酸作動性ニューロンとして同定された37).この交感神経プレモーターニューロン群は脊髄に軸索投射して褐色脂肪組織や皮膚血管を支配する交感神経節前ニューロンに対してグルタミン酸作動性シナプスを形成し(図3),一部のシナプスではグルタミン酸に加えてセロトニンも共放出する38, 39).脊髄のグルタミン酸作動性シナプスを阻害すると褐色脂肪熱産生が起こらなくなることから37),グルタミン酸が交感神経節前ニューロンを興奮させる主要な伝達物質だが,共放出されるセロトニンはグルタミン酸による神経興奮作用を増強すると考えられている40).
ラットに寒冷刺激や感染性発熱刺激(後述)を与えると,延髄縫線核でVGLUT3陽性の交感神経プレモーターニューロン群が活性化する37).また,寒冷刺激や感染性発熱刺激を与えることによって起こる褐色脂肪熱産生,ふるえ熱産生,皮膚血管収縮,脈拍上昇はすべて,神経活動抑制薬(ムシモールなど)を延髄縫線核に微量注入することによって完全に消失する6, 27, 41–43).したがって,延髄縫線核の交感神経プレモーターニューロン群は寒冷防御や感染性発熱のための体温調節反応を駆動する重要な役割を担う.ふるえ熱産生は交感神経ではなく体性運動神経を介して惹起されるが,延髄縫線核のVGLUT3陽性ニューロンは脊髄前角の運動ニューロンへも興奮性シグナルを入力してふるえ熱産生を惹起する可能性が考えられる.また,延髄縫線核のVGLUT3陽性ニューロンには心臓を支配する交感神経プレモーターニューロンも含まれていると考えられ,それらのニューロンを介して脈拍上昇が起こると考えられる(図3).
環境温度を徐々に低下させると,皮膚血管収縮,非ふるえ熱産生,ふるえ熱産生の順で惹起され,それぞれの生理反応が開始する環境温度閾値が異なる5).したがって,交感神経プレモーターニューロンや視床下部背内側部のニューロンには,それぞれの効果器に対応するサブグループが存在し,同じニューロンが複数の異なる効果器を支配することは少ないと考えられる(図3).
2)視床下部背内側部の駆動ニューロン
延髄縫線核の交感神経プレモーターニューロン群は,上位の視床下部背内側部から興奮性シグナルの入力を受けることで活性化し,交感神経系や運動神経系(ふるえ)を駆動する.たとえば,光遺伝学的手法を用いて視床下部背内側部から延髄縫線核へ至る神経伝達を選択的に刺激すると,褐色脂肪熱産生,脈拍,血圧が上昇する33).この興奮性投射ニューロンは視床下部背内側部の背側部に密集して局在し7, 33, 44),延髄縫線核のVGLUT3陽性交感神経プレモーターニューロンに対してVGLUT2含有のシナプス様構造を形成するグルタミン酸作動性ニューロンである33)(図3).また,ラットに寒冷刺激または感染性発熱刺激を与えると,視床下部背内側部から延髄縫線核へ軸索投射するニューロンが活性化される45, 46).さらに,寒冷刺激や感染性発熱刺激を与えることによって起こる褐色脂肪熱産生,ふるえ熱産生,脈拍・血圧上昇は,神経細胞抑制薬を視床下部背内側部に微量注入することによって消失する6, 7, 27, 47).しかし,寒冷刺激や感染性発熱刺激によって起こる皮膚血管収縮は,視床下部背内側部の神経興奮を阻害しても抑制されない48).したがって,視床下部背内側部から延髄縫線核へのグルタミン酸作動性の興奮性神経伝達は,寒冷環境や感染時における熱産生や循環器反応を駆動する上で必須である一方,皮膚血管収縮を介した熱放散抑制反応を駆動するものではなく,視索前野から視床下部背内側部を経ずに延髄縫線核の交感神経プレモーターニューロンへ至る直接路を通じて制御されると考えられる(図3).ただ,後述するように,視床下部背内側部のニューロンは心理ストレスによる皮膚血管収縮を駆動する働きがあることがわかっている.
また,視床下部背内側部が駆動する血圧上昇反応は,延髄縫線核の交感神経プレモーターニューロンと,血圧維持を担う吻側延髄腹外側野(rostral ventrolateral medulla:RVLM)の交感神経プレモーターニューロン49)の両方を介して起こると考えられている50, 51)(図3).しかし,視床下部背内側部から延髄縫線核への直接投射は知られているものの,視床下部背内側部からRVLMへの神経路については不明の点が多い.
上述したように,視床下部背内側部から延髄縫線核の交感神経プレモーターニューロン群への興奮性神経路については複数のサブグループがあり,それぞれのサブグループが異なる効果器を支配しているものと考えられる5).しかし,視床下部背内側部で効果器ごとのサブグループを同定するための選択的分子マーカーは知られていない.その一方で,視床下部背内側部で交感神経制御に関わるニューロンが発現する分子の解析は徐々に進んでいる.たとえば,視床下部背内側部から延髄縫線核へ投射するニューロンの中で,TrkBニューロトロフィン受容体を発現するものは褐色脂肪熱産生を促進する52).また,延髄縫線核へ投射するグルタミン酸作動性の視床下部背内側部の交感神経駆動ニューロンは,レプチン受容体(LepRb)やボンベシン受容体サブタイプ3(BRS3)を発現するサブグループを含む53, 54).これらのサブグループはともに,心拍数に影響を及ぼし,褐色脂肪熱産生を惹起できる54–56).一方,視床下部背内側部にはGABA作動性ニューロンのマーカーを発現するニューロン群も存在し,視床下部背内側部から延髄縫線核へ投射するTrkB発現ニューロンの約3分の1を占める52).視床下部背内側部ニューロンのさらなる分子解析により,多様な効果器を制御する交感神経駆動ニューロン群の各遺伝子マーカーが明らかになることが期待される.
3)視索前野の体温調節ニューロン
i. 体温調節の司令塔:EP3ニューロン
視床下部の最吻側部に位置する視索前野には体温調節中枢があり,そのシグナルが,下流の視床下部背内側部や延髄縫線核を経た興奮性出力を調節することで体温調節反応が制御される(図3~5).視索前野からの下行性の制御様式を示唆する興味深い実験がある.ラット視索前野のすぐ尾側の位置をナイフで切断し,視索前野からの下行性の神経伝達を遮断すると,褐色脂肪熱産生が異常に亢進し,著しい高体温状態に陥った57).ヒトでも視床下部の損傷によって高体温症が起こる症例が知られる58).このことは,視索前野が常に抑制シグナルを下流に出力し,それによって視床下部背内側部から延髄縫線核を経た興奮性神経路の暴走を抑制していることを示唆する.視索前野から出力されるこの抑制シグナルは深部体温のレベルを調節するために重要であり,そのシグナルを出力する視索前野ニューロンは脳の体温調節システムの司令塔の役割を担うと考えられる.最近,そのニューロンがラットで同定された34).
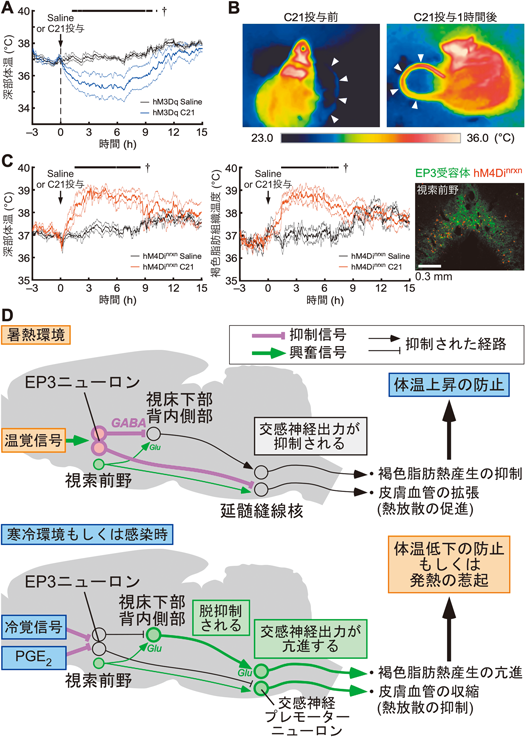
ラットを暑熱(36°C),寒冷(4°C)あるいは室温(25°C)に2時間曝露し,視索前野で活性化されるニューロンをFosタンパク質の発現を指標に探索すると,プロスタグランジンEP3受容体を発現するニューロン(EP3ニューロン)が暑熱曝露によって活性化されていた.一方,寒冷や室温への曝露後のFos発現は低レベルであった34).そこで,遺伝子改変ラットと化学遺伝学の手法を組み合わせて,視索前野のEP3ニューロンの活動を活性化すると,皮膚血管が拡張して積極的な熱放散反応が起こり,深部体温が35°C台にまで低下した(図6A, B).一方,EP3ニューロンの活動を抑制すると,褐色脂肪熱産生が惹起され,深部体温が約39°Cまで上昇した34)(図6C).視索前野のEP3ニューロンに膜移行型蛍光タンパク質を発現させて軸索終末まで可視化すると,視床下部背内側部や延髄縫線核など,さまざまな脳領域への軸索投射が観察され,視床下部背内側部へ投射した軸索の80,000個以上の終末を解析すると,その多くがGABA作動性シナプスのマーカーであるVGATを含有する一方,グルタミン酸作動性シナプスのマーカーであるVGLUT2はほとんど検出されなかった34).また,このVGAT含有軸索終末の中には,延髄縫線核へ投射する視床下部背内側部ニューロンにシナプス様の構造を形成するものが観察された.さらに,光遺伝学と組み合わせたスライスパッチクランプ記録を行い,視索前野のEP3ニューロン由来の軸索終末を光刺激すると,視床下部背内側部ニューロンでシナプス後電流が記録され,そのほとんどはGABA作動性シナプスを介したもので,グルタミン酸作動性シナプスを介した電流はわずかであった34).
このような実験結果から,視索前野のEP3ニューロンは常にGABA作動性の抑制シグナルを出力して,視床下部背内側部から延髄縫線核を経た交感神経駆動神経路を制御すると考えられる(図4, 6D).特に,EP3ニューロン群は二つの異なるサブグループが視床下部背内側部もしくは延髄縫線核へ軸索を投射し59),視床下部背内側部への投射は熱産生や脈拍・血圧を,延髄縫線核への投射は皮膚血管運動をそれぞれ制御すると考えられる.暑熱環境では外側腕傍核を経た温覚入力によってEP3ニューロンの活動が高まり,GABA作動性の抑制シグナルが強まるため,下流の交感神経駆動神経路が抑制され,熱産生が抑制されるとともに熱放散(皮膚血管拡張)が促進されることで体温の上昇を防ぐ.一方,寒冷環境では外側腕傍核を経た冷覚入力によってEP3ニューロンの活動が抑制され,EP3ニューロンからの抑制シグナルが弱まるため,下流の交感神経駆動神経路が脱抑制されて興奮し,熱産生が亢進するとともに皮膚血管が収縮して熱放散が抑制されることで体温の低下を防ぐ(図6D).視索前野のEP3ニューロンは体温調節中枢の司令塔の役割を担っており,EP3ニューロンから常時出力されるGABA作動性の抑制シグナルの強さが,調節される体温のレベルを決定していると考えられる.一方で,EP3ニューロンが深部体温(脳温)をモニターする温ニューロンであるかどうかは現時点では不明であり,今後解明すべき課題である.
また興味深いことに,ラットを34°Cの暑熱環境で2週間飼育すると,EP3ニューロンから視床下部背内側部へ投射する軸索のGABA作動性(VGAT含有)終末の数が有意に増加した34).この現象は,暑熱環境で視床下部背内側部の交感神経駆動ニューロンを効率的に抑制することによって,熱産生を強く抑圧する,暑熱順化のための変容の一つであると考えられる.今後,多様な温度環境に適応するためにEP3ニューロン群がどのように変容するのかを,形態だけでなく遺伝子発現などの変化もとらえながら解析することで,脳の生体調節システムの環境適応メカニズムが明らかになるかもしれない.
ii. 日内休眠時や睡眠時に体温を低下させるニューロン
近年,視索前野にはEP3ニューロン群以外にも体温を変動させる作用を持つニューロン群が報告されている.下垂体アデニル酸シクラーゼ活性化ポリペプチド(PACAP)と脳由来神経栄養因子(BDNF)を共発現する視索前野ニューロン(図4)を光遺伝学的に刺激したマウスでは,皮膚血管拡張と褐色脂肪熱産生抑制を伴う低体温と寒冷探索行動が誘導される60).このPACAP/BDNFニューロン群は皮膚の加温によって活性化されるが,温度感受性ニューロンのような温度応答性は示さない60).また,このニューロン群はGABA作動性とグルタミン酸作動性の両方のサブグループを含むが,視床下部背内側部へ投射するサブグループの90%以上はGABA作動性ニューロンのマーカーを発現し,その軸索終末を光遺伝学的に刺激すると褐色脂肪熱産生が抑制される60).特に,視索前野の腹側部のPACAPニューロンを刺激するとマウスの日内休眠と同様の著しい(室温レベルに至る)体温低下を誘導する一方,このニューロンの神経伝達機能を抑制しても体温は上昇しない61).同様に,pyroglutamylated RFamide peptide(QRFP)を発現する視索前野ニューロンを刺激しても著しい低体温が生じるが,その神経伝達機能を抑制しても体温は上昇しない62).PACAP/BDNFニューロン群やQRFPニューロン群は,一部がEP3ニューロン群と重複するものの62),体温を上昇させる機能がみられないことから,平常時の温熱恒常性に機能するものではなく,低温環境において空腹状態に陥った際に活性化され,日内休眠や冬眠様の状態を誘導する役割を担うものと考えられる.一方で,視索前野のEP3ニューロン群を活動操作すると体温を上下に変動させることができることから,EP3ニューロン群の主体は平常時の温熱恒常性に機能すると考えられる.
視索前野の中でEP3ニューロン,PACAP/BDNFニューロン,QRFPニューロンは正中視索前核や内側視索前野といった正中部に局在するが60–63),外側視索前野の腹側部のGABA作動性ニューロン群の一部も刺激すると体温が低下する64, 65).その体温低下作用は,視床下部背内側部ではなく延髄縫線核への直接のGABA作動性伝達を介した褐色脂肪熱産生抑制によるものであることが示されている65).また,このGABA作動性ニューロン群は,ガラニンを発現し,睡眠時の体温低下を誘導する働きを持つことが示唆されている66, 67)(図4).
iii. 内在性ニューロン
環境温度に応じたEP3ニューロンの活動変化は脊髄−外側腕傍核を経た温度感覚情報の入力に依存して起こると考えられるが,上述したように,外側腕傍核の冷覚と温覚を中継するニューロン群はともにグルタミン酸作動性の入力を視索前野へもたらす.したがって,外側腕傍核からの冷覚と温覚の興奮性シグナルはいったん,視索前野の正中視索前核の異なる内在性ニューロン群(視索前野内の別のニューロンに中継するニューロン群)に入力し,冷覚は抑制性(GABA作動性)の内在性ニューロン群の活性化を通じてEP3ニューロンを抑制する一方,温覚は興奮性(グルタミン酸作動性)の内在性ニューロン群の活性化を通じてEP3ニューロンの活動を高めると考えられる(図4).このモデルは,視索前野内への薬物の微量注入実験の結果によって支持されている26, 27).また,正中視索前核のNOS1陽性のグルタミン酸作動性の内在性ニューロンは内側視索前野や外側視索前野へ伝達するが,このニューロンを刺激すると,温覚入力時と同様に体温が低下する68).
iv. 興奮性投射ニューロン
冷覚シグナルが視索前野へ入力し,EP3ニューロンの活動が低下すると,視床下部背内側部のニューロンが抑制から解除(脱抑制)されるが,このとき,視床下部背内側部のニューロンが興奮するためには脱抑制に加えて興奮性の入力も必要である.視索前野からの下行性の出力をナイフで切断するだけで熱産生が亢進するという実験結果57)は,視索前野よりも尾側の他の脳領域から視床下部背内側部へ常時,交感神経駆動のための興奮性シグナルが入力されていることを示唆するが,現時点ではその脳領域は不明である(図4).最近では,視索前野からもグルタミン酸作動性ニューロンが興奮性シグナルを視床下部背内側部へ入力して褐色脂肪熱産生を駆動する可能性が示唆されており69),BRS3を発現するニューロン群である可能性が考えられる70).同様に,視索前野から延髄縫線核へもグルタミン酸作動性ニューロンの興奮性伝達があり,皮膚血管収縮を駆動する71, 72).視床下部背内側部や延髄縫線核の交感神経駆動ニューロンの活動は,視索前野のEP3ニューロンから入力されるGABA作動性の抑制シグナルと,視索前野やその他の領域から入力されるグルタミン酸作動性の興奮性入力とのバランスによって決定され,そのバランスの調節にはEP3ニューロン由来の抑制シグナルによる制御が大きな役割を担っていると考えられる(図4, 6D).
感染や全身性の炎症が起こると,寒冷刺激を受けたときと同様の熱産生反応や皮膚血管収縮が惹起され,体温が上昇する.これが発熱(fever)であるが,発熱は宿主にとって細菌感染を生き延びるために有益である73, 74)一方で,高熱が有害な場合もある75).発熱による体内温度の上昇は,病原体の活動至適温度を逸脱させることによってその増殖を抑制するとともに76),免疫細胞の活動を高める77)ことで宿主の防御力を高めると考えられる.
1)免疫–脳シグナル伝達メカニズム
発熱は感染や全身性の炎症の情報が免疫系から脳へ伝達されることによって起こる.しかし,脳の実質への物質透過は血液脳関門によって制限されるため,血中の情報伝達物質が脳実質内へ単純に拡散して神経回路に作用することは難しい.免疫–脳シグナル伝達における画期的な発見は,リポ多糖(LPS)をラットに全身投与する細菌感染モデルによって,脳の血管内皮細胞内でシクロオキシゲナーゼ-2(COX-2)やミクロソーム型プロスタグランジンE合成酵素(mPGES)などのプロスタグランジンE2(PGE2)生合成を担う酵素群が発現誘導されることを見いだしたことである78–80).この発見をもとに,感染や炎症に反応した免疫細胞から放出される炎症性サイトカイン(IL-1, IL-6, TNF-α)が脳の血管内皮細胞に作用し,血管内皮細胞内でCOX-2やmPGESを発現させ,その結果産生されるPGE2が脳実質内の神経回路に作用することで発熱が惹起されるという免疫–脳シグナル伝達メカニズム80)が提示された.このメカニズムの正しさは,マウス脳の血管内皮細胞選択的にCOX-2あるいはmPGESを欠損させると,LPS投与による発熱が強く抑制されるという実験結果81)によって確認された.脳の血管内皮細胞はLPS投与による長時間の発熱を誘発するPGE2の主要な供給源であるが,それに加えて,肺や肝臓のマクロファージもまた,末梢のPGE2供給源であることが示されている.末梢血中を循環するPGE2が何らかの仕組みを介して脳実質に拡散し,初期段階の発熱に関わると考えられている82, 83).
2)発熱誘発の中枢神経路メカニズム
このようにして産生されたPGE2は視索前野のEP3ニューロンに作用し,その神経活動を抑制することが発熱の引き金となる34).EP3受容体を介した神経活動の抑制作用は,この受容体が主に抑制性Gタンパク質Giと共役すること84)と矛盾しない.EP3受容体を視索前野選択的に欠損させたマウスでは感染性発熱が強く抑制されることから85),感染性発熱の誘発は視索前野のPGE2-EP3受容体シグナリングを介したメカニズムが主体である.この発熱メディエーター,PGE2を視索前野に微量注入すると,褐色脂肪熱産生,ふるえ熱産生,皮膚血管収縮,脈拍・血圧上昇が起こり,体温が上昇するが,その際に延髄縫線核の交感神経プレモーターニューロンが活性化される37, 41).視索前野へのPGE2投与によって起こるこれらの生理反応はすべて,神経活動抑制薬の微量注入によって延髄縫線核のニューロンの活動を抑制すると完全に消失することから27, 41, 42, 48, 86),延髄縫線核のプレモーターニューロンを経た遠心路シグナルが発熱の惹起に必須である.
延髄縫線核へ軸索投射する視床下部背内側部のニューロンはLPS投与による発熱誘発時に活性化し45),また,視索前野へのPGE2投与によって起こる褐色脂肪熱産生,ふるえ熱産生,脈拍・血圧上昇は視床下部背内側部へ神経活動抑制薬を微量注入することによって完全に消失する7, 27).これらのことから,感染時に産生されるPGE2が視索前野のEP3ニューロンに作用し,その神経活動が抑制されると,冷覚シグナルによる反応と同様に,視床下部背内側部から延髄縫線核への興奮性神経路が脱抑制され,熱産生や循環器反応が駆動されると考えられる.一方,PGE2投与によって起こる皮膚血管収縮は視索前野のEP3ニューロンから延髄縫線核への直接の抑制シグナルの減弱によって駆動されると考えられる48, 59)(図3,図6D).
これまで,感染性発熱は視索前野の体温調節中枢が決定する概念的な「セットポイント」の上昇によって起こると教科書的に説明されてきたが,現在では,PGE2の作用によって,視索前野のEP3ニューロンが出力する恒常的なGABA作動性の抑制シグナルのトーンが低下することがセットポイントの上昇をもたらすというモデルが示されている2, 34).これは,視索前野のEP3ニューロンの活動のトーンが体温のセットポイントを反映するという上述の考え方を支持するものである.
多くの心理ストレスは交感神経系を活性化し,血圧,脈拍,体温を上昇させる.これらの生理反応は,骨格筋や中枢神経系へ供給する血液(酸素と栄養)供給を増加させるとともにそれらの器官を温めることによって,たとえば野生動物が天敵と対峙するような場面などで身体パフォーマンスを向上させ,「闘争か逃走か」という状況を生き延びる上で有益であると考えられている.しかし,過度の,時に慢性的な心理ストレスは,ヒトにおいて異常な交感神経反応を伴う心身症を引き起こす.その症状の中には,心因性発熱(psychogenic fever)と呼ばれる慢性的な高体温症が知られ87),不明熱の約半数の症例が実は心因性発熱であったという報告もある88).心因性発熱の治療が困難なのは,PGE2産生を阻害する解熱剤に抵抗性を示すためであり,このことは感染性発熱とは異なる仕組みで生じる体温上昇であることを意味する89, 90).近年の研究から,心因性発熱(ストレス性体温上昇)の中枢神経メカニズムが明らかになってきた.
1)視床下部–延髄神経路
視床下部背内側部は「防衛領域(defense area)」として知られ,心理ストレスに対する多様な情動反応を惹起する中枢神経路のハブとして機能している91, 92).視床下部背内側部の背側部と腹側部にはそれぞれ,延髄縫線核あるいは視床下部室傍核(paraventricular hypothalamic nucleus)へ軸索投射するニューロン群が局在する33).これらのニューロン群はともに心理ストレスを受けた際に活性化され33, 45),延髄縫線核へ投射するニューロン群は交感神経反応を駆動する一方,視床下部室傍核へ投射するニューロン群は視床下部–下垂体–副腎皮質系を駆動してコルチコステロンなどのストレスホルモンを分泌させると考えられている33, 93)(図3).また,心理ストレスを受けた際には延髄縫線核のVGLUT3陽性交感神経プレモーターニューロンも活性化される90).
ラットやマウスを用いたストレス研究において,人間の心理社会ストレスを模倣したストレスモデルとして汎用される社会的敗北ストレスは,被験オス個体をより大きな支配的なオス個体と対峙させるストレスであり,社会的敗北ストレスを受けた個体は褐色脂肪熱産生や脈拍・血圧の上昇を示して体温が上昇する.これらのストレス性交感神経反応は,視床下部背内側部や延髄縫線核におけるニューロンの不活性化あるいはグルタミン酸受容体拮抗薬の注入によって強く抑制される33, 94).このことから,寒冷防御や感染性発熱と同様に,心理ストレス反応も視床下部背内側部から延髄縫線核の交感神経プレモーターニューロンへのグルタミン酸作動性の興奮伝達を介して駆動される(図5).ただし,血圧上昇の一部は視床下部背内側部からRVLMへ至る何らかの神経路が寄与する可能性がある50).
2)視床下部室傍核の寄与
視床下部室傍核はさまざまなストレスによって活性化するニューロン群が多く,ストレスホルモン分泌だけでなく,交感神経反応にも重要な部位であると考えられてきた.たとえば,視床下部室傍核からRVLMへの神経伝達を刺激すると交感神経性の心血管反応を惹起することができる95).しかし,ストレス性交感神経反応(心血管反応)に対する視床下部室傍核の寄与は限局的であると報告する論文も多い96).
一方,最近の研究で,視床下部室傍核のオキシトシン産生ニューロンが延髄縫線核の交感神経プレモーターニューロンを直接活性化して褐色脂肪熱産生や脈拍上昇を駆動することがわかった97).オキシトシン産生ニューロンは出産や授乳,子育て,交尾などの向社会行動や情動を伴う行動の際に活性化されることから98),視床下部室傍核から延髄縫線核へのオキシトシン神経伝達は,向社会行動や正の情動に伴う交感神経反応の惹起に関わる可能性が考えられる.また,このオキシトシン神経伝達による褐色脂肪熱産生(脂肪燃焼)促進は肥満の防止に働いている可能性がある97).
3)心身相関のメカニズム
心理ストレスによる交感神経反応の駆動の要となる視床下部背内側部へ心理ストレスシグナルを入力する上位の脳領域は世界中の研究者の興味の的であり,長年にわたって探索が行われてきた.これは,心理ストレスや情動を処理する皮質辺縁系の「心」の神経回路と,生体調節を担う視床下部の「体」の神経回路とをつなぐ「心身相関」の仕組みを解き明かすことにつながるからである.そして最近,ラットを用いた研究で心身相関の鍵となる神経路が発見された94).
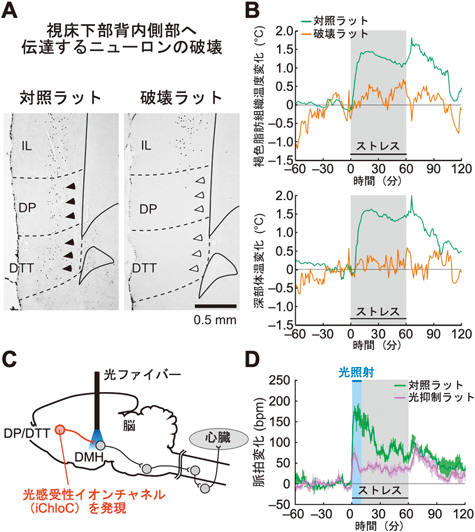
視床下部背内側部へ心理ストレスシグナルを入力する上流の神経路を探索する機能的神経トレーシング解析を行った結果,大脳皮質の内側前頭前皮質の最腹側部の背側脚皮質(dorsal peduncular cortex:DP)と背側蓋紐(dorsal tenia tecta:DTT)と呼ばれる,それまでほとんど研究が行われていなかった領域の錐体細胞が視床下部背内側部へ軸索投射し,心理ストレスシグナルを伝達することがわかった94).ラットに社会的敗北ストレスを与えるとDP/DTTの錐体細胞が活性化され,視床下部背内側部でVGLUT1陽性の軸索終末からグルタミン酸作動性のシナプス伝達を行うことで,延髄縫線核へ投射する交感神経駆動ニューロンを活性化する94).DP/DTTから視床下部背内側部への神経路を選択的に破壊あるいは伝達抑制すると,社会的敗北ストレスによる褐色脂肪熱産生,体温,脈拍,血圧の上昇が強く抑制された94)(図7).また興味深いことに,この神経路の伝達を抑制したラットは,ストレス源の個体から逃避する行動を示さなくなり,ストレスによる皮膚血管収縮反応も起こらなくなった94).この実験結果は,DP/DTTから視床下部背内側部への神経路が心理ストレス性交感神経反応だけでなくストレス逃避行動をも駆動することを意味する.上述したように,視床下部背内側部は寒冷刺激や感染性発熱刺激による皮膚血管収縮反応には関与しないが,心理ストレスによる皮膚血管収縮反応の駆動には関わり,熱放散の抑制を通じてストレス性体温上昇に寄与する(図3)94).加えて,DP/DTTへは視床や島皮質の一部などの皮質辺縁系を構成するニューロンから心理ストレスシグナルが入力される94).
こうした結果から,DP/DTTは皮質辺縁系のストレスシグナルを統合した上で,グルタミン酸作動性の興奮性マスターシグナルを視床下部背内側部へ入力することによってさまざまな交感神経反応だけでなくストレス逃避行動をも駆動することが明らかになった(図3).このように,DP/DTTから視床下部背内側部への神経路は,「心」と「体」の神経回路をつなぐことで心身相関を担う機能を持つ.重要なことに,この神経路を遮断しても平常時の体温調節や循環調節には影響しないことから94),ストレスシグナル伝達に特化した神経路である.このことから,この神経路は心因性発熱やパニック障害などのストレス関連疾患の症状を治療するための副作用の少ない創薬標的として有望視される.
DP/DTTの他に心理ストレス性体温上昇に関わることが報告されている脳領域として,不快情動やストレスに関わる外側手綱核(lateral habenula)があり,この脳部位を破壊すると心理ストレスによる褐色脂肪熱産生と体温上昇が減弱する99).しかし,外側手綱核から視床下部背内側部や延髄縫線核へ至る神経路は不明である99, 100).その他に,視床下部外側野のオレキシン産生ニューロンがストレス性体温上昇を修飾する作用が示されているが101),オレキシン産生ニューロンは脳の広範な領域に投射してさまざまな神経機能を修飾するため,どの領域でのオレキシン作用がストレス反応修飾に関わるかは明確ではない.
哺乳類が飢餓状態に陥ると生体内のエネルギー消費(代謝)を低下させる.その一つの反応として,褐色脂肪熱産生に代表される適応熱産生反応が抑制される.したがって,低温環境では体温を維持できず低体温に陥ることもある.これは,飢餓を生き延びるために生体に備わった飢餓反応の一つである.その他の飢餓反応としては,エネルギー摂取を促進するための食欲の亢進とそれに伴う食物探索行動の増加があげられる.近年,こうした飢餓反応を駆動する神経回路メカニズムは体温調節の神経回路と連関して機能することが明らかになり,その仕組みが少しずつみえてきた.
1)飢餓シグナルの発生機序
生体が飢餓状態を感知し,脳へ伝達する仕組みはいくつか知られるが,グレリン-ニューロペプチドY(NPY)系を介したシグナル伝達はその一つである.絶食状態の胃からグレリンというペプチドホルモンが内分泌され,血流に乗って視床下部の弓状核(arcuate nucleus)に到達し,NPY産生ニューロンを活性化する102).活性化されたNPY産生ニューロンはさまざまな領域に投射した軸索の終末からNPYを放出するが,その中でも視床下部室傍核へNPYが放出されると飢餓シグナルが生じ,代謝の低下と摂食量の増加が起こる103–105).興味深いことに,NPYの中枢作用は,全身代謝量の中で基礎代謝量を超える部分の適応的なエネルギー消費(寒冷環境での適応熱産生など)を抑制することにより,寒冷環境での体温低下を起こす106).また,視床下部室傍核でのNPY作用による代謝抑制には,褐色脂肪熱産生の抑制が寄与する107, 108).視床下部のNPY作用によって生じる飢餓シグナルが飢餓反応を惹起する神経回路メカニズムはまだ不明な点が多いが,近年,飢餓反応を駆動する延髄のニューロン群が同定された108).
2)飢餓反応を駆動する延髄のニューロン
ラットの視床下部室傍核へNPYを微量注入すると褐色脂肪熱産生が抑制されるが,延髄縫線核へGABAA受容体拮抗薬を注入することによって起こる褐色脂肪熱産生は抑制されないことが見いだされた108).このことは,視床下部室傍核へのNPY作用によって生じる飢餓シグナルが何らかの神経路を介して,延髄縫線核の交感神経プレモーターニューロンをGABA作動性の入力を通じて抑制することを示唆する.交感神経プレモーターニューロンへGABA作動性入力をもたらす神経細胞群を探索したところ,延髄の網様体にある中間網様核と小細胞網様核(intermediate and parvicellular reticular nuclei:IRt/PCRt)に分布することがわかった108).このGABA作動性ニューロン群は,視床下部にNPYを作用させると発火頻度が増加し,GABA作動性伝達を通じて延髄縫線核のVGLUT3陽性交感神経プレモーターニューロンを抑制することで,皮膚冷却や感染性発熱刺激による褐色脂肪熱産生や脈拍上昇を強く抑制する108).IRt/PCRtのニューロンの活動を抑制しておくと,視床下部室傍核へNPYを作用させることによる褐色脂肪熱産生の抑制が起こらなくなる108).こうした実験結果から,視床下部NPYの飢餓シグナルによって活性化されるIRt/PCRtのGABA作動性ニューロンが,延髄縫線核の交感神経プレモーターニューロンを抑制することによって褐色脂肪熱産生を抑制することが飢餓時の主要な代謝抑制メカニズムの一つであることがわかった(図3,図5).
興味深いことに,IRt/PCRtのニューロンを刺激すると褐色脂肪熱産生が抑制されるだけでなく,咀嚼や唾液分泌が起こり,摂食量が増加した108).IRt/PCRtには咀嚼筋の運動リズムを形成する神経回路が存在し109),延髄縫線核へ投射するIRt/PCRtのGABA作動性ニューロンは咀嚼運動ニューロンへも投射する108).したがって,IRt/PCRtのGABA作動性ニューロンは熱産生を抑制すると同時に,咀嚼のリズム形成にも関与して摂食(エネルギー摂取)を促進すると考えられる(図3).視床下部室傍核からIRt/PCRtへ飢餓信号を伝達する神経路についてはいくつかのモデル2, 110)が考えられているものの,いまだ特定されていない.
体温の中枢神経制御システムは,視床下部背内側部から延髄縫線核を経た興奮性の交感神経駆動路を,上位の視索前野のEP3ニューロンが常時抑制し,その抑制の強さを変化させることによって体温を制御する(図5,図6D).すべて興奮性神経路で構成するのではなく,恒常的な抑制シグナルによって興奮出力を常時制御するという様式はエネルギー的に高コストだと考えられるが,興奮性神経路のオーバーシュート(暴走)を抑えることができ,微細な体温の調節を実現する上ではより適切な仕組みであると考えられる.
視索前野のEP3ニューロンが出力する抑制シグナルの強さによって体温が平熱域に調節されると考えられるが,約37°Cという設定温度(セットポイント)を決定する分子メカニズムはいまだ不明である.おそらく,視索前野の温度感受性ニューロンが感知する深部体温の情報をもとにして,深部体温が約37°Cという設定温度に調節されるようにEP3ニューロンの発火頻度のゲインを制御する何らかの分子メカニズムが存在するものと考えられる.
現在,地球規模の気候変動によって熱中症が大きな医学的問題になりつつある.実験動物だけでなくヒトの体温調節メカニズムを解明し,体温調節システムが暑熱順化へ向けて変容するメカニズムを理解することによって,暑熱環境を生き抜くための技術の開発につながるものと考えられる.また,上述したように,ストレス性体温上昇をとらえる実験系が端緒となって心身相関の中枢メカニズムの解明に至ったが,今後,体温という計測が容易な生体パラメーターを利用することによって,心理や情動を処理する脳の神経回路メカニズムの解明や,さらには,さまざまなストレス関連疾患の根本的な発症メカニズムの解明と予防法の開発につながることが期待される.
加えて,脳の体温調節神経回路の加齢変容も取り組むべき研究課題である.たとえば,加齢によってヒトの褐色脂肪組織の熱産生能が低下することが見いだされ,それが脂肪の蓄積(中年太り)につながる可能性が指摘されている111).これは,脳の体温調節神経回路の加齢変容によって生じる可能性が考えられる.また,この神経回路の加齢変容は体温調節機能の低下をもたらし,高齢者における熱中症や低体温症の起こりやすさにつながる可能性がある.基礎生命科学と臨床医学の両面から体温調節研究を進める重要性がますます高まっている.