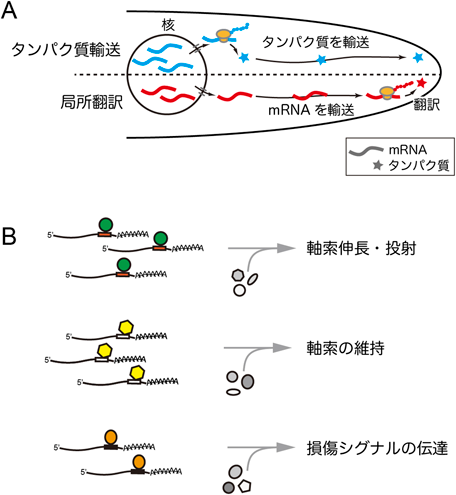
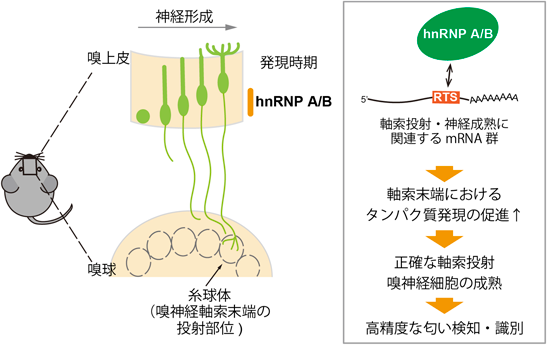
嗅神経回路の形成を担うRNA制御因子RNA binding proteins essential for olfactory circuit formation
新潟大学脳研究所動物資源開発研究分野Department of Comparative & Experimental Medicine, Brain Research Institute, Niigata University ◇ 〒951–8585 新潟県新潟市中央区旭町通1–757 ◇ 1–757 Asahimachidori, Chuo-ku Niigata, Niigata 951–8585, Japan
発行日:2024年2月25日Published: February 25, 2024