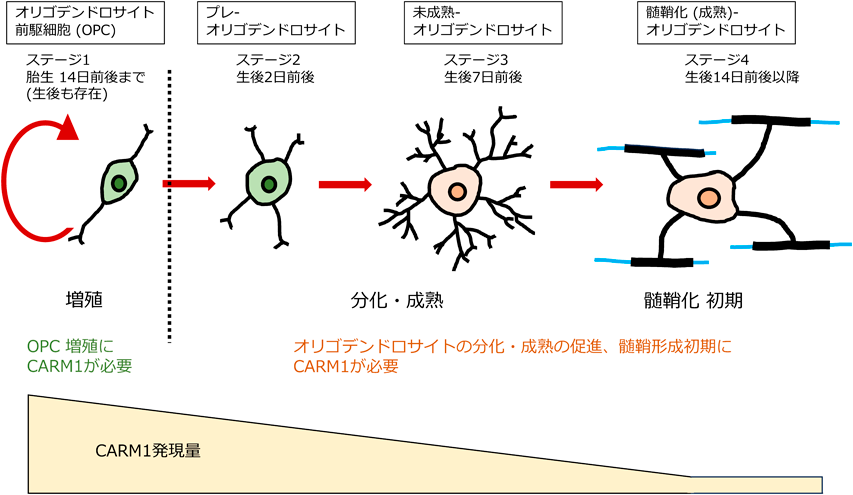
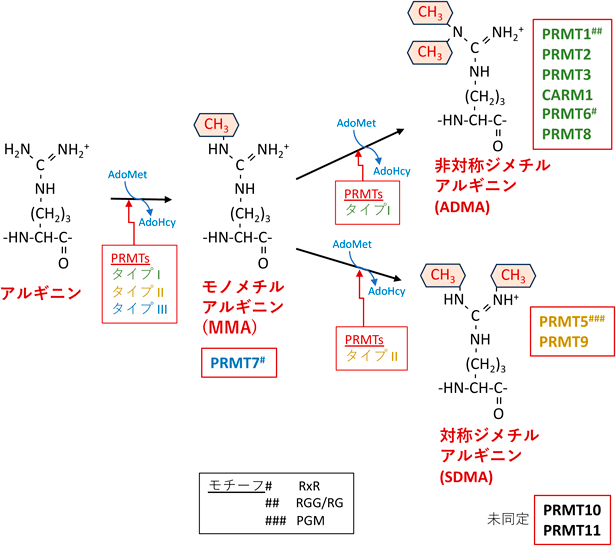
脳の発生および構造から見たアルギニン残基のメチル化制御Regulation of protein Arginine methylation during brain development
近畿大学東洋医学研究所分子脳科学研究部門Division of Molecular Brain Science, Research Institute of Traditional Asian Medicine, Kindai University ◇ 〒589–8511 大阪府大阪狭山市大野東377–2 ◇ 337–2 Ohno-Higashi, Osaka-Sayama, Osaka 589–8511, Japan
発行日:2024年2月25日Published: February 25, 2024