ラミノパチーにおける核膜の修復異常Defective nuclear envelope repair in laminopathies
金沢大学ナノ生命科学研究所WPI-NanoLSI, Kanazawa University ◇ 〒920–1192 石川県金沢市角間町ナノ生命科学研究所306 ◇ Kakuma-machi, Kanazawa, Ishikawa 920–1192, Japan
発行日:2024年2月25日Published: February 25, 2024
© 2024 公益社団法人日本生化学会© 2024 The Japanese Biochemical Society
真核細胞の核では,ゲノムDNAが核膜に包み込まれることによって細胞質から隔離されている.核膜は細胞分裂の際に崩壊し再構成されるが,それ以外の場合においても,細胞が押しつぶされたり,引っぱられたりするなどの機械的ストレスによって,核膜の一部が破壊されることが知られている1, 2).
ヒトを含む動物細胞では,核ラミナと呼ばれる核膜の内側を裏打ちする構造によって核の形態や機能を維持する役割がある.核ラミナの主要な構造タンパク質である核ラミン分子として,A型ラミン(ラミンA, C)とB型ラミン(ラミンB1, B2)が体細胞で広く発現している.ラミンAとCはLMNA遺伝子から転写・翻訳されたスプライシングバリアントであり,ラミンB1とB2はそれぞれLMNB1遺伝子とLMNB2遺伝子にコードされている.A型ラミンの遺伝子全長にわたって発見された500か所以上の変異は,心筋症,筋ジストロフィー,早老症を含む組織特異的な遺伝的疾患であるラミノパチーを発症する.
細胞の核膜が破壊された部位ではラミンB1が欠失し,ラミンAまたはCが局在することはすでによく知られている1, 3–6).しかし,ラミンAとCのいずれが局在しているのか,さらにこれらのラミンが核膜の修復メカニズムにどのように関与するのかはよくわかっていなかった.また,ラミンAとCは,これまで同一のタンパク質であるかのように扱われてきており,ラミンCに焦点を当てている研究は数少なかった.さらに,ラミンは核膜のタンパク質として知られているため,核質での役割については重要視されてこなかった.核質に豊富に局在するラミンCが核膜修復に寄与するという筆者らが見いだした新たな知見は,ラミノパチーの分子病態や,浸潤・転移するがん細胞と核ラミナの機能を理解するためにきわめて重要であると考えられる.
筆者らのグループは,共焦点蛍光顕微鏡の405 nmのレーザーを微小領域に照射することで細胞の核膜の一部を任意に破壊できる技術7)を用いて,核膜が破壊された直後におけるラミンの分子動態を解析した.レーザー微小照射によって細胞の核膜の一部を破壊してから10分後と60分後に各ラミン特異的抗体で免疫蛍光染色した結果,60分後のほとんどの細胞で破壊部位にラミンAとCが集積しているのに対し(ラミンA:~65%,ラミンC:100%),10分以内ではラミンCのみが集積していることがわかった(図1A)8).さらに,核膜破壊と迅速な修復過程を調べるために緑色蛍光タンパク質mEmeraldで標識したラミンを用いてライブセルイメージングを行ったところ,ラミンCのみが核膜が破壊された直後から迅速に集積すること(図1B)を見いだした8).
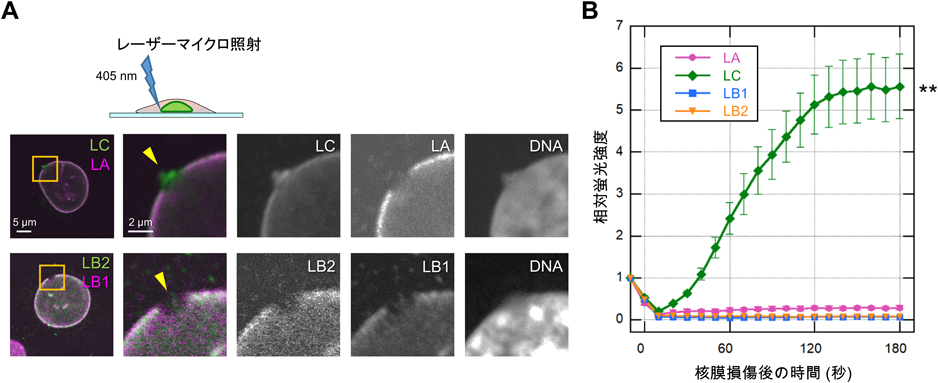
レーザー微小照射によって細胞の核膜の一部を破壊して,(A)10分後に各ラミン特異的抗体で免疫蛍光染色した.(B)緑色蛍光標識した各ラミンの破壊部位への集積を生きた細胞で経時的に解析した(文献8より改変).
なぜ各ラミン分子の中でラミンCのみが迅速に核膜破壊部位へ集積することができるのかを調べるために,ラミンCの詳細なドメイン解析およびラミノパチー変異体を作製し,レーザー微小照射による核膜破壊の修復動態への影響を解析した.まず,ラミンCはラミンAよりも核質に豊富に局在することが知られていることから,ラミンCをラミンA/Cノックアウト細胞で発現させ,核膜と核質の成分のうち,核質の成分を488 nmのレーザーで蛍光退色させてから核膜を破壊すると,破壊部位へのラミンCの集積が消失した8).さらに,核移行シグナル(NLS)を欠失させたラミンCでも同様に破壊部位への集積が消失したことから,ラミンCが破損部位へリクルートされるためにはNLSによって細胞質から核内に輸送され,核質にあらかじめ豊富に局在していることが必要であることを見いだした(図2)8).
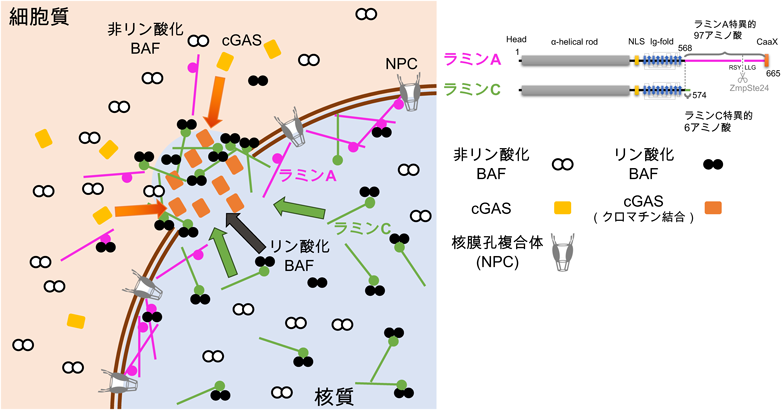
ラミンAとCは,両者に共通するヘッドドメイン,α-ヘリカルロッドドメイン,核移行シグナル(NLS),免疫グロブリン様ドメイン(Ig-fold)を含む568アミノ酸領域と,それぞれの特異的テール領域を有する.核膜の一部が破壊されるとラミンCとリン酸化BAFが核質から破壊部位へ迅速に集積する.細胞質のcGASは核から突出したクロマチンに集積する.NPC:nuclear pore complex.
Barrier-to-autointegration factor(BAF)は,細胞分裂期に核膜が再構築される際,核膜タンパク質の一部を染色体の表層にリクルートすることが知られているが,間期の核膜修復においても一部の核膜内膜タンパク質を集積させることが報告された7).ラミンA/Cも免疫グロブリン様(Ig-fold)ドメインを介してBAFと結合し9),核膜再構築の際にリクルートされることから10),核膜修復の際にもBAFによって破壊部位に集積することが考えられる.そこで,ラミノパチー変異のうちラミンCのNLS以降のIg-foldドメインを含むテール部位の欠失変異体と,BAFとの結合に影響があると報告されているIg-fold変異体を解析したところ,両者において破壊部位への迅速な集積が消失または有意に減弱した8).さらに,BAFのノックダウンでもラミンCの集積が有意に減弱したため,BAFがIg-foldドメインと結合することがラミンCの集積に重要であることを見いだした(図2)8).
BAFの核局在にラミンA/Cが関与していることはすでに報告されていたが,どのラミンが重要であるのか不明であった11).筆者らはまず野生型,ラミンA/C, B1, B2の各ノックアウト細胞を用いてBAFの核局在を免疫染色によって調べたところ,ラミンA/Cノックアウト細胞において核局在が消失していることを確認した.また,細胞質のBAFは核内よりもリン酸化が少ないことが知られているため12),ウエスタンブロットによって発現量を調べたところ,リン酸化BAFのタンパク質発現量が減少していることを新たに見いだした8).さらに,ラミンAとCのいずれか,あるいは両方がBAFの核局在に関与しているのかを調べるために,それぞれのラミン特異的なshRNAを用いてノックダウンした細胞を解析すると,ラミンA, CノックダウンのどちらもBAFの核局在が消失し,ウエスタンブロットの結果,ラミンAとCのノックダウンによって相加的なリン酸化BAFの減少がみられたことから,BAFの核局在にはラミンAおよびCの両者が関与していることが明らかとなった8).
核膜破損によってDNAが核内から細胞質に突出すると,DNAセンサーであるcyclic GMP-AMP synthase(cGAS)がDNAと結合する1, 2, 7).cGASは細胞質内に侵入した外来の二本鎖DNAに結合すると活性化し,cyclic GMP-AMP(cGAMP)というシグナル伝達物質を合成することによって,stimulator of interferon gene(STING)を介した自然免疫を活性化する酵素として知られている.自己の染色体では,cGASがDNAではなくヌクレオソームと選択的に結合することで不活性化されている13–15).
筆者らはまず,赤色蛍光タンパク質sfCherryで標識したcGASを一過性に細胞に発現させたところ,核と細胞質に局在する細胞がみられたため,それぞれのcGAS局在の細胞でレーザー微小照射によって細胞の核膜の一部を破壊してライブセルイメージングをを行った.その結果,cGASが細胞質局在している細胞ではBAFとラミンCが同時に集積し,少し遅れてcGASが集積するのに対し,cGASが核局在している細胞では破損部位へのcGASの集積がみられなかった8).これら核と細胞質に局在するcGASをそれぞれfluorescence recovery after photobleaching(FRAP)により解析したところ,細胞質のcGASは退色させてから直ちに蛍光が回復するのに対し,核局在のcGASは退色させてから6分後でもほとんど回復しないことから,ヌクレオソームと結合していることが示唆された.興味深いことに,ヒトとマウスのcGASではヌクレオソーム結合サイトの数が異なることが知られているが16),FRAPではヒトとマウスどちらも同様の結果が得られた.
また,cGASとBAFはDNAへの結合で競合すると報告されているが17),cGASが核局在して破損部位へ集積しない細胞ではBAFとラミンCの集積が減弱しており,超解像顕微鏡を用いてラミンC, BAF, cGASの局在を調べたところ,BAFは核から突出した染色体の先端に集積しているのに対し,cGASはBAFとラミンCが集積している位置よりも内側のDNAに集積していることが明らかとなった8).
さらに,野生型および各ラミンノックアウト細胞でレーザー微小照射によって細胞の核膜の一部を破壊してから10分後にBAF抗体とcGAS抗体で免疫染色した結果,野生型とラミンB1, B2の各ノックアウト細胞のほとんどの細胞でBAFとcGASが破壊部位に集積しているのに対し,ラミンA/Cノックアウト細胞ではどちらも集積が減弱していることがわかった(図3).同様の結果がライブセルイメージングでも確認できたことから,核から突出した染色体にBAFが結合すること,ならびに損傷したDNAを感知するcGASがDNAと結合することの双方に,ラミンAまたはCが関与していること(図3)が明らかとなった.
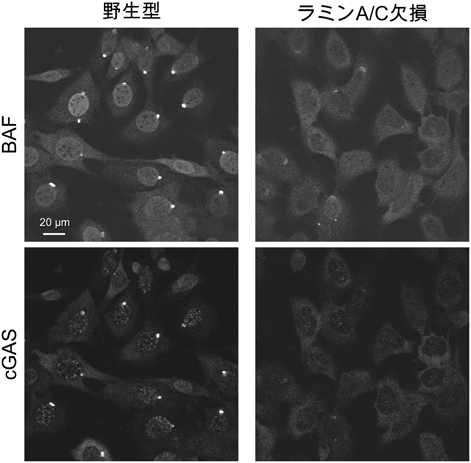
ラミンA/C欠損細胞において核膜が破壊されてから10分後のBAFとcGASの破壊部位への集積が減弱した(文献8より改変).
筆者らは,硬く線維化した組織のすき間を浸潤するがん細胞などが圧縮されたり,筋肉細胞などが強く伸縮したりすることによって,核膜の一部が破壊されるという現象から着想を得て,その核膜の修復機構へのラミンの関与について詳細に解析した.その結果,各ラミン特異的抗体やラミンA,ラミンC特異的ノックダウンなどの手法を用いることで,ラミンAとCが機能的に異なるということを示した革新的な研究となった.本稿で引用している論文でも,単にラミンAと表記されているが実際にはラミンA/Cの両方を標的としているものや,表記の紛らわしい抗体製品も多く存在するため注意が必要である.またB型ラミンは遺伝子発現の耐容量が小さく,過剰発現すると核膜に異常を来すが,ラミンAは過剰発現させると核質への局在が増加し,ラミンCに似た核膜への集積動態を示すため,低い発現量に抑えることが必要である.しかし,このラミンAの過剰発現によるアーティファクトであっても核膜を破壊したときの集積動態はラミンAとCの間で大きな差がみられたため8),ラミンA特異的テール領域に核膜への迅速な集積を阻害する分子が結合している可能性があると考えられる.本研究では,ラミンCをノックダウンすると,核膜を破壊したときの核質成分の細胞質への漏出が加速することを明らかにした.さらに,Ig-foldドメイン内にラミノパシー変異が入るとBAFと結合できないために核膜の初期修復に遅延が生じることが明らかとなった8).これまで機械的ストレスなどによる核膜の破壊において核膜の修復機構へのラミンの関与が示唆されており,本研究で修復初期にどのラミンが集積するかを特定したことは,細胞生理学的な意義がきわめて大きいと考えられる.本研究成果から,破壊された核膜の修復機構,および核膜の恒常性の維持機構の解明につながるだけでなく,今後は心筋症,筋ジストロフィー,早老症を原因するラミノパシー変異について着目し,分子レベルでの病態解明や新規治療戦略の探索などの応用研究へと展開していくと期待される.
本研究はJSPS科研費JP20KK0158, JP20K06617, JP18H05527, JP21H04764の助成および文部科学省先端研究基盤共用促進事業(コアファシリティ構築支援プログラム)JPMXS0440200021で共用された機器を利用した成果です.また本研究を遂行するにあたり,東京工業大学オープンファシリティーセンター,ライフサイエンス推進機器共同利用室,基礎研究機構に多大なご支援をいただきました.ここに感謝いたします.最後に,本稿の執筆にあたりご協力いただきました「文部科学省世界トップレベル研究拠点プログラム(WPI)」に深く感謝申し上げます.
1) Denais, C.M., Gilbert, R.M., Isermann, P., McGregor, A.L., te Lindert, M., Weigelin, B., Davidson, P.M., Friedl, P., Wolf, K., & Lammerding, J. (2016) Nuclear envelope rupture and repair during cancer cell migration. Science, 352, 353–358.
2) Raab, M., Gentili, M., de Belly, H., Thiam, H.R., Vargas, P., Jimenez, A.J., Lautenschlaeger, F., Voituriez, R., Lennon-Duménil, A.M., Manel, N., et al. (2016) ESCRT III repairs nuclear envelope ruptures during cell migration to limit DNA damage and cell death. Science, 352, 359–362.
3) Harada, T., Swift, J., Irianto, J., Shin, J.W., Spinler, K.R., Athirasala, A., Diegmiller, R., Dingal, P.C., Ivanovska, I.L., & Discher, D.E. (2014) Nuclear lamin stiffness is a barrier to 3D migration, but softness can limit survival. J. Cell Biol., 204, 669–682.
4) Xia, Y., Pfeifer, C.R., Zhu, K., Irianto, J., Liu, D., Pannell, K., Chen, E.J., Dooling, L.J., Tobin, M.P., Wang, M., et al. (2019) Rescue of DNA damage after constricted migration reveals a mechano-regulated threshold for cell cycle. J. Cell Biol., 218, 2545–2563.
5) Young, A.M., Gunn, A.L., & Hatch, E.M. (2020) BAF facilitates interphase nuclear membrane repair through recruitment of nuclear transmembrane proteins. Mol. Biol. Cell, 31, 1551–1560.
6) Sears, R.M. & Roux, K.J. (2022) Mechanisms of A-type lamin targeting to nuclear ruptures are disrupted in LMNA- and BANF1-associated progerias. Cells, 11, 865.
7) Halfmann, C.T., Sears, R.M., Katiyar, A., Busselman, B.W., Aman, L.K., Zhang, Q., O’Bryan, C.S., Angelini, T.E., Lele, T.P., & Roux, K.J. (2019) Repair of nuclear ruptures requires barrier-to-autointegration factor. J. Cell Biol., 218, 2136–2149.
8) Kono, Y., Adam, S.A., Sato, Y., Reddy, K.L., Zheng, Y., Medalia, O., Goldman, R.D., Kimura, H., & Shimi, T. (2022) Nucleoplasmic lamin C rapidly accumulates at sites of nuclear envelope rupture with BAF and cGAS. J. Cell Biol., 221, e202201024.
9) Samson, C., Petitalot, A., Celli, F., Herrada, I., Ropars, V., Le Du, M.H., Nhiri, N., Jacquet, E., Arteni, A.A., Buendia, B., et al. (2018) Structural analysis of the ternary complex between lamin A/C, BAF and emerin identifies an interface disrupted in autosomal recessive progeroid diseases. Nucleic Acids Res., 46, 10460–10473.
10) Haraguchi, T., Kojidani, T., Koujin, T., Shimi, T., Osakada, H., Mori, C., Yamamoto, A., & Hiraoka, Y. (2008) Live cell imaging and electron microscopy reveal dynamic processes of BAF-directed nuclear envelope assembly. J. Cell Sci., 121, 2540–2554.
11) Lin, Q., Yu, B., Wang, X., Zhu, S., Zhao, G., Jia, M., Huang, F., Xu, N., Ren, H., Jiang, Q., et al. (2020) K6-linked SUMOylation of BAF regulates nuclear integrity and DNA replication in mammalian cells. Proc. Natl. Acad. Sci. USA, 117, 10378–10387.
12) Zhuang, X., Semenova, E., Maric, D., & Craigie, R. (2014) Dephosphorylation of barrier-to-autointegration factor by protein phosphatase 4 and its role in cell mitosis. J. Biol. Chem., 289, 1119–1127.
13) Kujirai, T., Zierhut, C., Takizawa, Y., Kim, R., Negishi, L., Uruma, N., Hirai, S., Funabiki, H., & Kurumizaka, H. (2020) Structural basis for the inhibition of cGAS by nucleosomes. Science, 370, 455–458.
14) Zhao, B., Xu, P., Rowlett, C.M., Jing, T., Shinde, O., Lei, Y., West, A.P., Liu, W.R., & Li, P. (2020) The molecular basis of tight nuclear tethering and inactivation of cGAS. Nature, 587, 673–677.
15) Michalski, S., de Oliveira Mann, C.C., Stafford, C.A., Witte, G., Bartho, J., Lammens, K., Hornung, V., & Hopfner, K.P. (2020) Structural basis for sequestration and autoinhibition of cGAS by chromatin. Nature, 587, 678–682.
16)鯨井智也,胡桃坂仁志(2021)自然免疫DNAセンサーcGASが自己の染色体DNAによって不活性化されるメカニズム.生物物理,61, 324–326.
17) Guey, B., Wischnewski, M., Decout, A., Makasheva, K., Kaynak, M., Sakar, M.S., Fierz, B., & Ablasser, A. (2020) BAF restricts cGAS on nuclear DNA to prevent innate immune activation. Science, 369, 823–828.

金沢大学ナノ生命科学研究所 研究員.
1987年大阪市に生る.2011年鳥取大学医学部生命科学科卒業.13年同大学院医学系研究科機能再生医科学専攻博士前期課程修了(再生医科学).17年カノンキュア株式会社(鳥取大学発ベンチャー)研究員兼代表取締役.18年鳥取大学大学院医学系研究科助教を経て,20年東京工業大学細胞制御工学研究センター研究員.23年より現職.
研究テーマと抱負再生医療学,核ラミン,核膜破壊,プロジェリア,ラミノパチーの分子病態の解明と治療法の開発研究.
ウェブサイトhttp://researchmap.jp/yoheikono
趣味旅行,ドライブ,鉄道,野球,ゴルフ,スノボ,漫画,アニメ,映画,ラジオ,免許・資格取得(大型一種,中型二種,大型特殊,けん引,大型二輪,乗馬ライセンス4級,車両系建設機械(整地),フォークリフト,小型移動式クレーン,玉掛け).

金沢大学ナノ生命科学研究所 特任准教授.博士(理学).
2005年大阪大学大学院理学研究科生物科学専攻博士後期課程修了.同年情報通信研究機構未来ICT研究所特別研究員.06年米国ノースウエスタン大学博士研究員.12年同助教.15年米国シカゴ大学上級研究員.16年米国ノースウエスタン大学助教.18年東京工業大学特任准教授を経て,23年より現職.
研究テーマと抱負細胞核の構造と機能,核ラミナ,核ラミン,ラミノパチーの発症機序の解明と治療法の開発.
ウェブサイトhttps://researchmap.jp/shimi
趣味音楽,料理.

This page was created on 2024-01-17T12:00:42.149+09:00
This page was last modified on 2024-02-16T13:56:29.000+09:00
このサイトは(株)国際文献社によって運用されています。