糖鎖ハイブリッドナノマテリアルの設計と機能Design and function of glycan Hybrid nanomaterials
京都大学大学院医学研究科Graduate School of Medicine, Kyoto University ◇ 〒606–8501 京都府京都市左京区吉田近衛町 ◇ Yoshida Konoe-cho, Sakyo-ku, Kyoto 606–8501, Japan
発行日:2024年4月25日Published: April 25, 2024
筆者は糖鎖ハイブリッドの自己組織化制御(集合,集積制御)による糖鎖ナノマテリアルの設計,および,糖鎖加水分解酵素を利用したアミロース糖鎖ハイブリッドの設計とバイオ応用を展開してきた.コレステロール基をプルランに導入した疎水化多糖(CHP)は安定なナノゲル(粒径30 nm)を形成し,分子シャペロン機能を持つため,内部に取り込んだタンパク質を安定化する.この特性を利用して,がん免疫ワクチンとしてがん免疫治療応用を,また,カチオン性CHPナノゲルは経鼻ワクチンとして感染症予防応用を展開し,実用化を含めた研究が進められている.本稿では,グルカンホスホリラーゼによって糖鎖プライマーの非還元末端から糖鎖伸長が可能であることや,プライマーの還元末端の化学修飾が可能なことを利用した,さまざまなアミロースハイブリッドの設計とそのバイオ応用に関する研究を紹介する.
© 2024 公益社団法人日本生化学会© 2024 The Japanese Biochemical Society
糖鎖(多糖)は,地球上で生み出されるバイオ資源であり,材料(高機能先端材料,汎用材料),食糧・食品素材,創薬などに活用されている.持続可能な開発目標(sustainable development goals:SDGs)への貢献においても,重要なマテリアル資源である.一方,タンパク質や核酸のようにいわゆるセントラルドグマに直接組み込まれていないこともあり,合成,構造・機能解析は,いまだ課題が残されている.糖鎖(多糖)は,単純なグルコースの重合体であるセルロースやアミロースにおいてさえも,高次構造やナノ構造などの構造多様性がある.この化学構造から由来する糖鎖(多糖)の物性・機能の多様性こそが糖鎖の魅力であり,材料素材としてのポテンシャルの高さを示している.近年,有機化学,高分子科学,自己組織科学が著しく進展し,糖鎖(多糖)の精密合成,集積制御,機能解析の体系化によって生み出されるグライコマテリアル素材群は,環境・産業・バイオ・医療など幅広い分野へと波及している.
革新的な次世代ナノ医療において,核酸,タンパク質などのバイオ医薬品の開発とその実用化が期待されている.バイオ医薬品は一般に安定性が低く製剤化が難しいことや体内での分解や不活性化を受けやすく半減期が非常に短いなどの問題もあり,安定に目的の部位,かつ必要な時間に,送達あるいは徐放させることのできるドラッグデリバリーシステム(Drug Delivery System:DDS)を可能とする再生医療用材料の進展が必要不可欠となっている.そのため,この問題を解決しうるバイオマテリアル開発は重要であり,バイオマテリアル素材の一つとして,生体適合性に優れた糖鎖(多糖)が注目されている.抗体をはじめとするタンパク質への糖鎖の付加による安定性や指向性の制御,糖鎖導入DDSナノキャリア,多糖ゲルによる再生医療用材料などの研究が進んでいる.多糖の持つ構造や機能の多様性に加えて,その生体親和性・分解性の高さから糖鎖を機能性素子として用いた糖鎖ハイブリッド材料の開発が行われている.本稿では,糖鎖を創る,操作するという観点から,糖鎖ハイブリッドナノ材料の設計と機能について,我々の研究を紹介する.
水酸基を多く有する高分子である多糖は,アルギン酸や寒天(アガロースとアガロペクチン)など,ヒドロゲル(水を含んだゲル)を形成する特性を有しており,食品をはじめさまざまな用途に用いられている.一般に高分子ゲルは,高分子鎖間が部分的に架橋された三次元ネットワーク(網目)構造を有していることが特徴であり,その内部にさまざまな物質を閉じ込めることができる.架橋点が共有結合である化学架橋ゲルや静電的相互作用,水素結合,van der Waals力や疎水性相互作用などの分子間力で形成される物理架橋ゲルが知られている.生体系では可逆的なゲル形成が可能な物理架橋ゲルが多い.生体の多糖やタンパク質から形成される細胞外マトリクスもゲル構造を有しており,サイトカインや細胞外小胞などの情報伝達物質を貯蔵,徐放して細胞間情報を制御することで,細胞の組織化による器官,臓器形成を助けている.
我々は,生体適合性の高い多糖を利用したバイオマテリアル研究を図ってきた.水溶性多糖に部分的に疎水基を導入した疎水化多糖を設計し水中での会合挙動を解析したところ,疎水基が多糖の会合を誘起して30 nm程度のナノ微粒子を形成することを見いだした1, 2).生体系には,ガングリオシド,リポ多糖(LPS)などの糖脂質が存在し,生体膜への挿入や自身でのミセル形成など水中で自己組織的に会合する.我々の設計した疎水化多糖は,広い意味では糖脂質の仲間に入るものの,通常の糖脂質よりも糖鎖あたりの疎水基が圧倒的に少ない(50から100多糖あたり一つの疎水基)構造特性を有している.したがって,水中での会合体は,糖鎖がループ構造によりつながり,疎水基の会合領域を架橋点とするゲル構造を有することを明らかにした.特にプルラン(分子量10万,約600単糖)に,100単糖あたり1個程度でコレステロール基を疎水基として化学結合させたコレステロール置換プルラン(cholesterol-bearing pullulan:CHP)は,水中で安定な会合体ナノ微粒子,ナノゲルを形成することを見いだした(図1).いわゆる手にとれる通常のゲルをナノサイズまでダウンサイジングしたゲルである.さらに,我々はプルラン以外のさまざまな水溶性多糖(マンナン,デキストリン,サイクロアミロース,ヒアルロン酸,ヒドロプロピルセルロース,キシログルカン,グルカンデンドリマー)を用いた疎水化多糖ナノゲルライブラリーを構築している3).
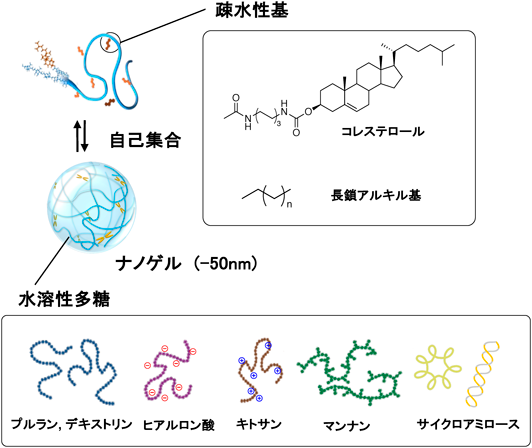
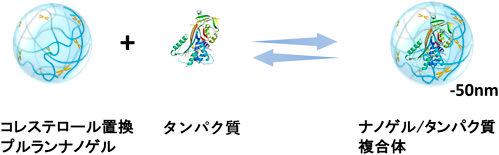
CHPナノゲルは,コレステロール基が約4分子程度会合した架橋点が複数存在するナノサイズのネットワークを有した物理架橋ゲルである.ナノ微粒子内の水含量が80%程度であり,ナノ微粒子内に疎水性のドメイン(架橋点)と親水性糖鎖ネットワークを有する両親媒性ゲルである.この構造特性による疎水的性質を主な駆動力として水溶性タンパク質を取り込むことを明らかにした.たとえば,CHPナノゲルとBSA(bovine serum albumin)は1対1のモル比で複合化しているが,複合化後のナノ微粒子のサイズは30 nm程度であり,複合化前後で粒子サイズがほとんど変わらないことからも,BSAはナノゲル内部に取り込まれていると考えられる(図2).また,分子量の小さなインスリンではナノゲルあたり9分子が複合化された.これらの結果から,CHPナノゲルは,分子量の比較的大きなタンパク質からペプチドレベルの小さなものまで効率よく取り込む高分子ホストであることを見いだした.タンパク質の疎水面とナノゲルの疎水性コレステロール基との相互作用,そして糖鎖のナノサイズネットワーク構造により,内包させたタンパク質はナノゲル内に安定に取り込まれていると考えられる4).
興味深いことに,CHPナノゲルは,まるで天然の分子シャペロンであるかのように振る舞い,部分的に変性して疎水面を露出した変性タンパク質を認識し,これをナノゲル内に選択的に取り込む.また,他のタンパク質の添加による交換反応や,コレステロールを包接するシクロデキストリンを添加することによるナノゲルの崩壊反応により,取り込んだタンパク質を天然状態へとリフォールディングしながら取り出すことができる.ナノゲルは,水溶液中でのタンパク質の加熱による変性凝集体を複合化することで抑制する,いわゆるヒートショックプロテイン機能を示す5).また,化学変性したタンパク質のリフォールディングを促進する6).in vitroの実験系において,アルツハイマー病の原因と考えられているAβタンパク質の凝集はナノゲル存在下で抑制され,その毒性が軽減したことより,ナノゲルには分子シャペロン様の機能もあることが示された7).
以上のように多糖にわずかに疎水基を導入するだけで,ナノゲル形成能と分子シャペロン機能という従来にない特性を有する多糖ナノ微粒子を構築しうることが明らかになった.自己組織化因子として,疎水基に限らず,静電的相互作用を利用したカチオン性多糖とアニオン性多糖からなるポリイオンコンプレックスナノゲル,オリゴ核酸どうしの相補的二重鎖形成を利用したオリゴ核酸多糖ナノゲルも設計した3, 8).また,ポリプロピレンオキシド(polypropylene oxide:PPO)は温度変化により水に対する溶解性が劇的に変化する温度応答性高分子材料であり,低温では水に溶解するが,下限臨界溶液温度(lower critical solution temperature:LCST)まで温度が上がると不溶化する性質をもつ.熱応答性のPPOをプルランに部分的に導入(グラフト化)した会合性多糖を用いると,LCST以上(5°C以上)でPPOの会合に伴う多糖高分子鎖のフォールディングが起こり,PPO二分子膜構造を有する単分散な熱可逆的高分子ベシクルが形成されることを明らかにした9).
以上のように,多糖の疎水化による自己組織化により,多糖の新たな機能を引き出す方法論を提案した.さらに,自己組織化ナノゲルをボトムアップ的に組み上げてナノゲルを架橋点とするさまざまな機能化ゲル材料の構築手法(ナノゲルテクトニクス)を確立し,足場ゲル材料として再生医療への応用も図っている3, 10–12).
分子シャペロン機能を有する疎水化多糖ナノゲル基盤材料を用いて,新規DDS技術や再生医療用バイオマテリアルの開発に取り組むことで応用研究を図ってきた.我々は,1990年代に,がん抗原タンパク質やペプチドを複合化したCHPナノゲルを用いたワクチンを開発した.抗原提示細胞に抗原タンパク質を運ぶナノキャリアとしての利用例である.ナノゲルワクチンは,抗原タンパク質の投与だけでは実現できない,がん免疫治療で重要なキラーT細胞を効率的に誘導しうる優れたがん治療用ワクチンであることを見いだした13).現在,ヒト臨床試験も含めた実用化に向けた研究が進められている.さらに,最近では,CHPナノゲルが糖鎖の受容体を介して,腫瘍関連マクロファージに選択的に取り込まれることを見いだし,免疫療法抵抗性のがんの治療成績の向上を実現できる可能性を見いだしている14).従来の治療法では効果のない難治性がんの克服に向けて,CHPナノゲルを用いた新しいがん免疫療法に関する研究が進められている.
がんワクチンは基本的には治療用のワクチンであるが,疎水化多糖ナノゲルを用いた予防ワクチンの開発も行っている.たとえば,新型コロナウイルス感染症(COVID-19)に対して現在広く用いられているmRNAワクチンよりも,リコンビナントタンパク質ワクチンは,より副作用の少ないワクチンとして期待されている.CHPナノゲルにCOVID-19のスパイクタンパク質を複合化したワクチンの感染予防における有効性は,マウス皮下投与実験により明らかにされた(投稿中).また,多糖として非イオン性のプルランの代わりに,アニオン性のヒアルロン酸を用いたコレステロール置換ヒアルロン酸ナノゲル15)を基盤としたワクチンは,凍結乾燥による保存も可能になるだけでなく,長期の免疫記憶保持の可能性を期待しうる優れた特性を持つことが最近明らかになっている.
これまで述べてきた皮下投与のワクチンとは違って,鼻粘膜に投与する経鼻ワクチンは粘膜での感染自体を予防できる次世代ワクチンとして注目されている.抗原タンパク質を複合化したカチオン性CHPナノゲルは,経鼻投与で効果的な予防ワクチンとして機能しうることが明らかになった16).抗原タンパク質を粘膜下の免疫細胞系に効率よく運ぶナノキャリアとしての利用である.たとえば,肺炎の原因として知られる肺炎球菌の予防ワクチンへ応用し,その有効性が明らかになっている17).現在,千葉大学未来粘膜ワクチン研究開発シナジー拠点の中で,新規なワクチンモダリティーとしてカチオン性CHPナノゲルを用いたさまざまな経鼻ワクチンの開発が進められている18).
生体系では,いくつかの酵素を用いてさまざまな多糖が合成されている.一方,試験管内合成では,糖加水分解酵素の可逆的酵素反応性を利用して,条件を選ぶことで加水分解の逆反応を進行させ,プライマーとなる糖鎖への糖転移を起こすことで,さまざまなオリゴ糖や多糖を合成しうることも比較的古くから知られている19).グリコーゲンホスホリラーゼ(glycogen phosphorylase:GP)を用いた人工アミロースの合成や合成ポリマーとの複合材料の開発20, 21),短鎖のアミロースから2種類の酵素(branching enzyme, amylomaltase)による単分散な分岐性球状ナノ粒子(グルカンデンドリマー)の合成22)とその機能化23),また,最近ではcellulaseやcellodextrin phosphorylaseを用いた合成セルロハイブリッド24)などが報告されている.以下では,我々が開発したさまざまなアミロースハイブリッドの設計と機能解析について報告する25).
アミロースは4~5糖のオリゴ糖をプライマー,グルコース1-リン酸を基質として,GPによる酵素重合から合成できる(図3).この合成反応はアミロースの非還元末端側から進行することから,プライマーの還元末端側を化学修飾した糖鎖ハイブリッドを用いて種々のアミロースハイブリッドを合成することができる.我々はマルトペンタオースの還元末端にポリエチレングリコール(PEG)を導入して酵素重合を行うことでPEGアミロースブロックポリマーを合成した26).PEGはクロロホルムに溶解するために,このブロックポリマーは水のみならずクロロホルムにも溶解し,特に少量の水を含むクロロホルム中ではアミロース鎖が水相に集合した逆相ミセルを形成した.直鎖状α(1,4)グルカンであるアミロースは,疎水性化合物の存在下でヘリックスを形成して,そのヘリックス内部に疎水性化合物を取り込む“高分子ホスト分子”として知られている.PEGアミロースは逆相ミセル中で,アミロースの集積による特異な包接機能を示した27).このブロック型のPEGアミロースプライマーを拡張して,両末端2本鎖,さらに,4本鎖,8本鎖のPEGを有するプライマーを設計し,酵素重合することで多分岐アミロースを合成した.特に,8本鎖スターアミロース(各糖鎖重合度:約60)を有するアミローススターポリマー(糖鎖重合度:約60)は,その多価性によりアミロース単体では示さない低分子包接挙動や協同的な分子包接を示すことが明らかになった28).また,スペルミンを修飾したカチオン性8本鎖スターアミロースは,血管内皮細胞増殖因子の発現を抑制するsiRNAと複合化し,安定なナノ微粒子を形成するため,優れた核酸デリバリーシステムの構築につながる可能性が明らかになった28).
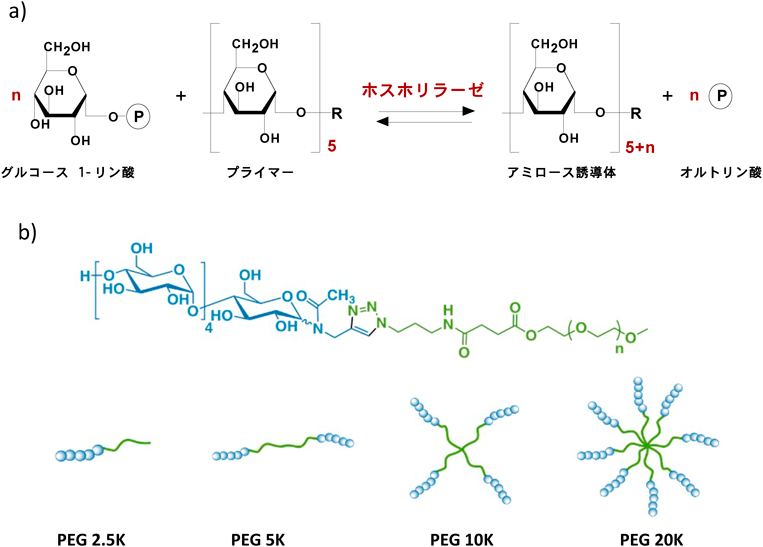
PEG 5K, 10K, 20Kが2本鎖,4本鎖,8本鎖PEGに相当する.
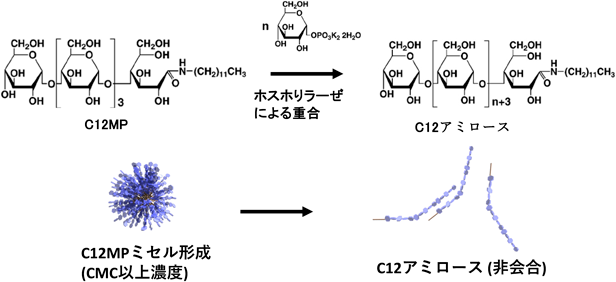
マルトペンタオースの還元末端にアルキル基を導入した両親媒性プライマーを合成した.ドデシル基を有するアルキルマルトペンタオース(C12MP)は,臨界ミセル濃度(critical micelle concentration:CMC)が0.5 mMである球状ミセルを形成した.このミセル溶液(1 mM)に,グルコース1-リン酸とAMPを加えて,筋肉GPによる重合反応を行ったところ,プライマー上のマルトペンタオースに7残基のグルコースが転移され,グルコース鎖の鎖長が約12残基に伸長されたときにミセルの崩壊がみられた(図4)29).12残基からなる糖鎖が付加されたことで球状ミセルのCMCが上昇し崩壊したものと考えられる.これにより酵素反応と共役した糖鎖界面活性剤の会合制御が可能となった.
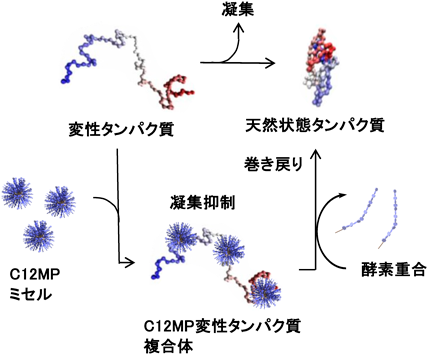
界面活性剤とシクロデキストリンを利用したタンパク質リフォールディングに対する人工分子シャペロン機能が報告されている30).まず,界面活性剤は疎水的会合力によって変性中間体と複合体を形成し,タンパク質の凝集を抑制し可溶化状態を維持する.その後シクロデキストリンを添加して界面活性剤を包接することでタンパク質を解離させ,それとともにタンパク質リフォールディングが進行するという2段階の分子シャペロンシステムである.
我々は,先に述べたアルキルマルトペンタオース(C12MP)ミセルの形成・崩壊の酵素反応制御系を利用した酵素応答性人工分子シャペロン系を設計した31).炭酸脱水酵素(carbonic anhydrase B:CAB)をモデルタンパク質として,化学変性剤である塩酸グアニジンによって変性させたCABをC12MPミセルによって巻き戻すことができるかどうかについての実験を行った.30 mg/mL CABを6 Mグアニジン塩酸塩を含む緩衝液(pH 7.5)中で室温16時間静置することで変性させた.この変性CABを1000倍希釈することでリフォールディングを行った.リフォールディング率は,天然状態のCABの酵素活性を100%としたときの酵素活性の回復率として表した.C12MPミセルが存在しない緩衝液系では,その回復率は20%前後で多くのタンパク質が凝集したままであった.一方,100 mMグルコース1-リン酸,10 mM AMPを含むC12MPミセル(1 mM)水溶液で変性CABを希釈すると凝集はほぼ抑制された.この水溶液に40°Cで筋肉GPを加えて重合を開始し,CABと複合化したC12MP分子上で糖鎖の伸長反応を行い親水性を増大させることで酵素複合体からのCABの解離と酵素活性の回復を調べた.GPを加えてから2時間後に酵素の活性回復率は65%を超え,C12MPミセルが人工分子シャペロンとして機能することが明らかになった(図5).酵素の代わりに従来の方法であるシクロデキストリンを添加した系では,2時間後に32%,24時間で57%しか回復率は上昇せず,酵素応答性人工分子シャペロンの方が,より迅速なリフォールディングシステムを構築できることが明らかになった.
リン脂質—界面活性剤系における酵素応答性ミセル-ベシクル転移について検討した.リン脂質とC12MPを混合すると動的光散乱測定より粒径20 nmの混合ミセルが形成することがわかった.この溶液に酵素処理を行うと時間とともに会合体の粒径が増加し,重合時間が2時間のとき(重合度14),比較的サイズのそろったリポソーム100 nmが形成された.これは,リポソーム形成法として従来にない新しい手法である29).
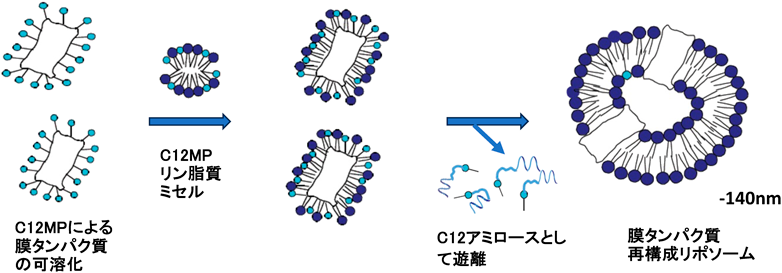
糖鎖界面活性剤は膜タンパク質の可溶化剤として利用されている.一般に,膜タンパク質再構成リポソームの調製には,まず糖鎖界面活性剤とリン脂質からなる混合ミセルで可溶化した膜タンパク質の複合体を形成させ,その後リポソームへ再構成させるときには,糖鎖界面活性剤だけを透析などで除去することでリポソームが自発的に形成(セル-ベシクル転移)され,それと同時に膜タンパク質再構成リポソームが得られる.そこで我々は,酵素重合によるミセル-ベシクル転移系を膜タンパク質の再構成系に適用することを考えた.ロドプシンを膜タンパク質モデルとして用い,まずC12MPミセルによりロドプシンを可溶化した.このロドプシン(2.5 µM)を含むミセル溶液(1 mM)にC12MP(5 mM)-リン脂質(DPPC)(5 mM)混合ミセルを加えた膜タンパク質ミセル複合体を形成させた.この溶液に対して,先と同様に酵素重合を行った.重合2時間後に280 nmのリポソームが形成した(図6).この再構成リポソームのロドプシンのプロトンポンプ活性を調べた.435 nmの波長のロングパスフィルターを通した光を照射することでpH値が上昇するとともに,光照射を停止するとpH値が急激に低下した.混合ミセル系では,そのような顕著なpH変化は認められないことから,ロドプシンがリポソームに効率よく再構成されていることが明らかになった29).本システムが,新規な膜タンパク質再構成リポソーム調製法として有効であることが明らかになった.
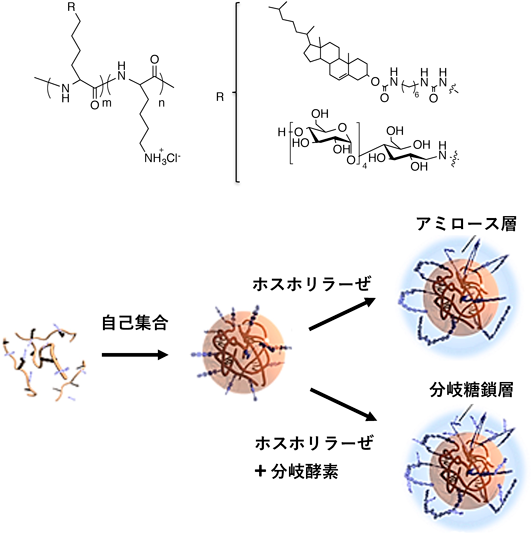
コレステロール基を置換した疎水化ポリリシンにマルトペンタオースを導入した疎水化糖鎖ポリペプチド(リシン100残基あたりコレステロール3.9個または10個,マルトペンタオース約30個を含む)が糖鎖ペプチドナノゲルを形成すること,さらに筋肉GPを用いた前述と同様な酵素重合を行うことでアミロース被覆ナノゲルを構築しうることを報告した32).このナノゲルをDDSに応用する場合には,糖鎖被覆によりナノゲルの表面電荷が遮蔽されるものの,アミロース鎖間の二重鎖形成に伴う二次凝集体形成により,ナノゲルの水中でのコロイド不安定性が問題となった.
この問題を解決するために,GPに加えて糖鎖分岐酵素であるglycogen branching enzyme(GBE)を用いて疎水化糖鎖ポリペプチドナノゲルに対してタンデム酵素重合を行い,多くの分岐を有する多糖を合成させることにより多分岐糖鎖ナノゲルを構築した(糖鎖の重合度15~20)(図7)33).ナノゲルを取り巻く糖鎖が分岐することで,老化による沈殿が抑制され,水中での安定性の高いナノゲルが得られた.また,糖鎖の被覆によりポリリシンの細胞毒性が軽減されることも明らかになった.このナノゲルはカチオン性のポリリシンの内部空間を有していることからsiRNAを複合化可能であり,さらに,糖鎖の被覆により血液成分の存在下でも複合体の安定性は維持された.がんの進行に関与する血管内皮細胞増殖因子であるVEGFの発現を抑制するsiRNAを用いた細胞実験により,VEGF遺伝子が効率よくノックダウンされ,優れたsiRNAデリバリーシステムとして機能することが明らかになった33).
1) Akiyoshi, K., Deguchi, S., Moriguchi, N., Yamaguchi, S., & Sunamoto, J. (1993) Self-aggregates of hydrophobized polysaccharides in water. Formation and characteristics of nanoparticles. Macromolecules, 26, 3062–3068.
2) Akiyoshi, K., Deguchi, S., Tajima, H., Nishikawa, T., & Sunamoto, J. (1997) Microscopic structure and thermoresponsiveness of a hydrogel nanoparticle by self-assembly of a hydrophobized polysaccharide. Macromolecules, 30, 857–861.
3) Hashimoto, Y., Mukai, S., Sasaki, Y., & Akiyoshi, K. (2018) Nanogel tectonics for tissue engineering: Protein delivery systems with nanogel chaperones. Adv. Healthc. Mater., 23, 1800729.
4) Nishikawa, T., Akiyoshi, K., & Sunamoto, J. (1996) Macromolecular complexation between bovine serum albumin and the self-assembled hydrogel nanoparticle of hydrophobized polysaccharides. J. Am. Chem. Soc., 18, 6110–6115.
5) Akiyoshi, K., Sasaki, Y., & Sunamoto, J. (1999) Molecular chaperone-like activity of hydrogel nanoparticles of hydrophobized pullulan: Thermal stabilization with refolding of carbonic anhydrase B. Bioconjug. Chem., 10, 321–324.
6) Nomura, Y., Ikeda, M., Yamaguchi, N., Aoyama, Y., & Akiyoshi, K. (2003) Protein refolding assisted by self-assembled nanogels as novel artificial molecular chaperone. FEBS Lett., 553, 271–276.
7) Boridy, S., Takahashi, H., Akiyoshi, K., & Maysinger, D. (2009) The binding of pullulan modified cholesteryl nanogels to Abeta oligomers and their suppression of cytotoxicity. Biomaterials, 30, 5583–5591.
8) Sasaki, Y. & Akiyoshi, K. (2010) Nanogel engineering for new nanobiomaterials: From chaperoning engineering to biomedical applications. Chem. Rec., 10, 366–376.
9) Nishimura, T., Shishi, S., Sasaki, Y., & Akiyoshi, K. (2020) Thermoresponsive polysaccharide graft polymer vesicles with tunable size and structural memory. J. Am. Chem. Soc., 142, 11784–11790.
10) Hashimoto, Y., Mukai, S., Sawada, S., Sasaki, Y., & Akiyoshi, K. (2015) Nanogel tectonic porous gel loading biologics, nanocarriers, and cells for advanced scaffold. Biomaterials, 37, 107–115.
11) Nakai, K., Yamamoto, K., Kishida, T., Kotani, S., Sato, Y., Horiguchi, S., Yamanobe, H., Adachi, T., Boschetto, F., Marin, E., et al. (2021) Osteogenic response to polysaccharide nanogel sheets of human fibroblasts after conversion into functional osteoblasts by direct phenotypic cell reprogramming. Front. Bioeng. Biotechnol., 9, 713932.
12) Hayashi, S., Sasaki, Y., Kubo, H., Sawada, S., Kinoshita, N., Marukawa, E., Harada, H., & Akiyoshi, K. (2021) Construction of hybrid cell spheroids using cell-sized cross-linked nanogel microspheres as an artificial extracellular matrix. ACS Appl. Bio Mater., 4, 7848–7855.
13) Tahara, Y. & Akiyoshi, K. (2015) Current advances in self-assembled nanogel delivery systems for immunotherapy. Adv. Drug Deliv. Rev., 95, 65–76.
14) Muraoka, D., Harada, N., Shiku, H., & Akiyoshi, K. (2022) Self-assembled polysaccharide nanogel delivery system for overcoming tumor immune resistance. J. Control. Release, 347, 175–182.
15) Nakai, T., Hirakura, T., Sakurai, Y., Shimoboji, T., Ishigai, M., & Akiyoshi, K. (2012) Injectable hydrogel for sustained protein release by salt-induced association of hyaluronic acid nanogel. Macromol. Biosci., 12, 475–483.
16) Nochi, T., Yuki, Y., Takahashi, H., Sawada, S., Mejima, M., Kohda, T., Harada, N., Kong, G., Sato, A., Kataoka, N., et al. (2010) Nanogel antigenic protein-delivery system for adjuvant-free intranasal vaccines. Nat. Mater., 9, 572–578.
17) Yuki, Y., Uchida, Y., Sawada, S., Ouchida-Nakahashi, R., Sugiura, K., Mori, H., Yamanoue, T., Machita, T., Honma, A., Kurokawa, S., et al. (2021) Characterization and specification of a trivalent protein-based pneumococcal vaccine formulation using an adjuvant-free nanogel nasal delivery system. Mol. Pharm., 18, 1582–1592.
18) Umemoto, S., Nakahashi-Ouchida, R., Yuki, Y., Kurokawa, S., Machita, T., Uchida, Y., Mori, H., Yamanoue, T., Shibata, T., Sawada, S., et al. (2023) Cationic-nanogel nasal vaccine containing the ectodomain of RSV-small hydrophobic protein induces protective immunity in rodents. NPJ Vaccines, 8, 106, 10.1038/s41541-023-00700-3.
19) Kadokawa, J. (2011) Precision polysaccharide synthesis catalyzed by enzymes. Chem. Rev., 111, 4308–4345.
20) Braumuhl, V., Jonas, G., & Stadler, R. (1995). Enzymic grafting of amylose from poly(dimethylsiloxanes). Macromolecules, 28, 17–24.
21) Kadokawa, J. (2022) Glucan phosphorylase-catalyzed enzymatic synthesis of unnatural oligosaccharides and polysaccharides using nonnative substrates. Polym. J., 54, 413–426.
22) Kajiura, H., Takata, H., Akiyama, T., Kakutani, R., Furuyashiki, T., Kojima, I., Harui, T., & Kuriki, K. (2011) In vitro synthesis of glycogen: The structure, properties, and physiological function of enzymatically-synthesized glycogen. Biologia (Bratisl.), 66, 387–394.
23) Takahashi, H., Sawada, S., & Akiyoshi, K. (2011) Amphiphilic polysaccharide nanoballs: A new building block for nanogel biomedical engineering and artificial chaperones. ACS Nano, 5, 337–345.
24) Sakurai, Y., Sawada, T., & Serizawa, T. (2022) Phosphorylase-catalyzed synthesis and self-assembled structures of cellulose oligomers in the presence of protein denaturants. Polym. J., 54, 561–569.
25) Nishimura, T. & Akiyoshi, K. (2017) Amylose engineering: Phosphorylase-catalyzed polymerization of functional saccharide primers for glycobiomaterials. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol., 9, e1423.
26) Akiyoshi, K., Kohara, M., Ito, K., Kitamura, S., & Sunamoto, J. (1999) Enzymatic synthesis and characterization of amphiphilic block copolymers of poly(ethylene oxide) and amylose. Macromol. Rapid Commun., 20, 112–115.
27) Akiyoshi, K., Maruichi, N., Kohara, M., & Kitamura, S. (2002) Amphiphilic block copolymer with a molecular recognition site: Induction of a novel binding characteristic of amylose by self-assembly of poly(ethylene oxide)-block-amylose in chloroform. Biomacromolecules, 3, 3280–3283.
28) Nishimura, T., Mukai, S., Sawada, S., & Akiyoshi, K. (2015) Glyco star polymers as helical multivalent host and biofunctional nano-platform. ACS Macro Lett., 4, 367–371.
29) Morimoto, N., Ogino, N., Narita, T., Kitamura, S., & Akiyoshi, K. (2007) Enzyme-responsive molecular assembly system with amylose-primer surfactants. J. Am. Chem. Soc., 129, 458–459.
30) Nishimura, T. & Akiyoshi, K. (2020) Artificial molecular chaperone systems for proteins, nucleic acids, and synthetic molecules. Bioconjug. Chem., 31, 1259–1267.
31) Morimoto, N., Ogino, N., Narita, T., & Akiyoshi, K. (2009) Enzyme-responsive artificial chaperone system with amphiphilic amylose primer. J. Biotechnol., 140, 246–249.
32) Morimoto, N., Yamazaki, M., Tamada, J., & Akiyoshi, K. (2013) Polysaccharide-hair cationic polypeptide nanogels: Self-assembly and enzymatic polymerization of amylose primer-modified cholesteryl poly(L-lysine). Langmuir, 29, 7509–7514.
33) Nishimura, T., Yamada, A., Umezaki, K., Sawada, S., Mukai, S., Sasaki, Y., & Akiyoshi, K. (2017) Self-assembled polypeptide nanogels with enzymatically transformable surface as a small interfering RNA delivery platform. Biomacromolecules, 18, 3913–3923.

京都大学大学院医学研究科 特任教授.工学博士.
1985年九州大学大学院工学研究科博士課程修了,米国パデュー大学博士研究員,87年長崎大学工学部講師,89年京都大学工学部助手,93年同大学院工学研究科助教授,99年JSTさきがけ研究員兼任,2002年東京医科歯科大学教授,10年京都大学大学院工学研究科教授,11年JST-ERATO研究総括兼任,23年より現職.
研究テーマと抱負バイオインスパイアード科学,ナノゲル工学,糖鎖工学,リポソーム工学,細胞外小胞工学,DDS・再生医療応用.
ウェブサイトhttps://kdb.iimc.kyoto-u.ac.jp/profile_private/ja.42378f75a40b4eb4.html
趣味草花散策,ドライブ,美味しい食事とワインを楽しむこと.

This page was created on 2024-03-19T17:46:28.228+09:00
This page was last modified on 2024-04-16T11:57:08.000+09:00
このサイトは(株)国際文献社によって運用されています。