ミトコンドリア由来小胞,古くて新しい膜輸送Mitochondrial-derived vesicles, the new classical membrane traffic
順天堂大学大学院医学研究科Juntendo University ◇ 〒113–8421 東京都文京区本郷2–1–1 ◇ 2–1–1 Hongo, Bunkyo-ku, Tokyo 113–8421, Japan
発行日:2024年4月25日Published: April 25, 2024
ミトコンドリアは多彩な機能を持つダイナミックな細胞小器官(オルガネラ)である.分裂や融合,伸長などの形態変化に加え,他のオルガネラとの接着,細胞内局在変化,細胞間移動などその動態も多様である.ミトコンドリアの機能維持において正常な動態制御は必須であるが,それぞれの動態は独立しているわけではなく,連関しながら恒常性が保たれている.ミトコンドリア由来小胞(mitochondrial-derived vesicles:MDVs)はミトコンドリアからカーゴ特異的に形成・放出される小胞で,ペルオキシソームやリソソームなどの細胞内膜構造に輸送される.多くの細菌が細胞外小胞を放出するようにMDVsは進化的にも保存された膜動態であるとも考えられるが,研究報告例も少なく,多くの謎に包まれたままである.本稿では近年徐々に明らかになってきたMDVsの機能や生理的意義について概説する.
© 2024 公益社団法人日本生化学会© 2024 The Japanese Biochemical Society
ミトコンドリアは細胞内エネルギー産生や鉄硫黄の形成などの代謝機能に加え,細胞死や自然免疫のシグナル伝達など多彩な機能を持つ細胞小器官(オルガネラ)である.先天的なミトコンドリアの機能異常はミトコンドリア病を引き起こすが,加齢やストレスによるミトコンドリアの機能低下が心疾患や糖尿病,神経変性疾患などに関わっていることが明らかになってきており,ミトコンドリアの機能維持は我々の健康や寿命においても重要である1, 2).ミトコンドリアは細胞内外の環境に応じて,分裂や融合を繰り返しながら形態や局在を変化させる非常に動的なオルガネラである.これらの動態制御因子の遺伝子異常が種々の疾患の原因となっているように,動態と機能は密接に関わっており,ミトコンドリアの機能維持において正常な動態制御も必須である3).また,近年のイメージング技術の飛躍的な向上により,ミトコンドリアが小胞体などの他のオルガネラと能動的に接着点を形成し,代謝物やシグナルの交換や形態制御の場として機能することが明らかになっており,ミトコンドリア動態の理解にはオルガネラネットワークの包括的な理解が必要となってきた.さらに,動態の範囲は細胞外へも飛び出し,トンネルナノチューブや小胞を介して細胞間を移動することも明らかとなり,ミトコンドリアは膜動態の面からも非常に魅力的なオルガネラである4).本稿では最初の報告から15年あまりが経過し,近年徐々に報告例も増えてきたミトコンドリア由来小胞(mitochondrial-derived vesicles:MDVs)について,発見の経緯やこれまでの研究,今後の展望について概説する.
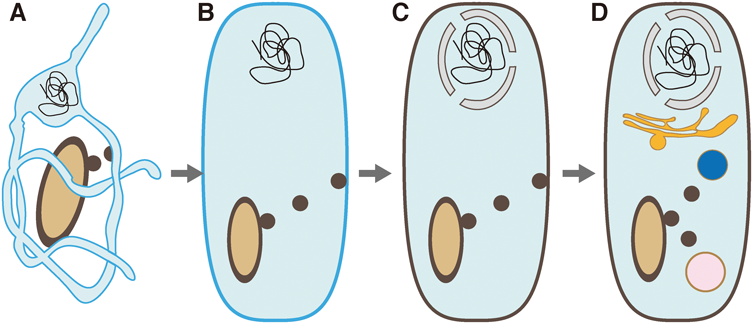
カナダ・オタワ大学のMcBride博士(現マギル大学)らのグループにより,世界で初めてミトコンドリアから小胞が形成・放出されることが報告された5).最初の報告ではMDVsがペルオキシソームへと輸送されることを報告しているが,その後の解析によりリソソームへの輸送経路も明らかとなった(図1)6).ペルオキシソームは最初のMDVs発見のきっかけとなった経路であるが,誘導条件も含めその生理的意義に関してはいまだに知られていない部分が多く残されている.一方,リソソームへの経路はストレス下で誘導されることなどから,酸化や変性タンパク質を分解するための品質管理機構であることが複数のグループより報告されている.これまでに,神経変性疾患7–9),心疾患10, 11),がんなどの疾患に加え12),老化細胞においてもMDVsとの関わりが報告されており13),細胞生物学的,生理的意義が徐々に明らかになってきている.
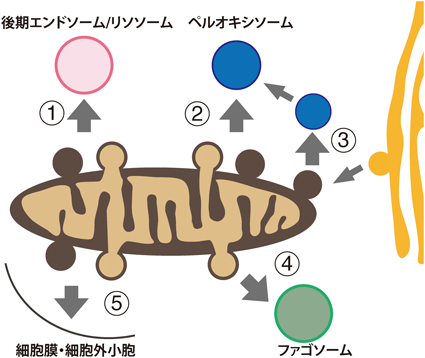
これまでに報告されているMDVsの種類と輸送先.本稿では左上後期エンドソーム/リソソームより番号順に時計回りで紹介していく.②は既存ペルオキシソーム,③はペルオキシソームde novo合成.
ミトコンドリアは核とは異なる独自のDNAを有していることなどから,共生説が広く受け入れられており,グラム陰性細菌であるアルファプロテオバクテリア由来であると考えられている.細菌は毒物などを小胞として細胞外へ放出していることからMDVsは自然な膜動態であるとも考えられるが14),ミトコンドリアから小胞が出るということは長らく一般的には受け入れられてこなかった.その理由としては,ミトコンドリアは分裂することが広く知られていることから,分裂して断片化したミトコンドリア,あるいは過剰発現系による人工産物であると理解されていたと考えられる.そこで,発見者であるMcBride博士らのグループは分裂ミトコンドリアと区別するために,次の三つの判断基準を提唱した15).
生物種・細胞種による多様性はあるが,ミトコンドリアの直径は500 nm~1 µmとされている.透過型電子顕微鏡で観察されるMDVsの直径は70~150 nmであり他の細胞内小胞と同程度である.ミトコンドリア外膜とMDVsの脂質成分は異なっており,MDVsの方がホスファチジン酸の含有率が高いことから膜の曲率や小胞構造の形成に寄与していることが示唆されている16).
ミトコンドリアを複数のマーカーで標識すると大部分はミトコンドリアネットワーク,あるいは断片化されて小さくなったミトコンドリアが共標識される.一方,あるマーカーでは標識されるが,別のマーカーでは標識されないような小胞様の構造も観察される(図2).これらはカーゴ選択的にミトコンドリアから放出されたMDVsであると考えられるが,非対称分裂した断片化ミトコンドリアである可能性もあるので,上記1)の条件と合わせて判断される17, 18).マーカーとしてはミトコンドリア外膜のタンパク質透過装置のサブユニットであるTOMM20とマトリックス内の代謝酵素であるPDH(pyruvate dehydrogenase)がよく用いられる.どちらもそれぞれ独立したMDVsあるいは二重陽性のMDVsとして放出される(図2).
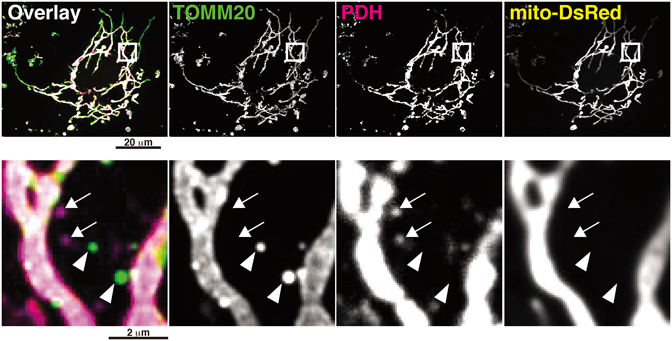
DRP1のドミナントネガティブ体とmito-DsRedを発現させたCOS-7細胞を固定後,抗TOMM20, PDH抗体を用いて免疫蛍光染色を行った.矢頭はTOMM20陽性/PDH陰性,矢印はTOMM20陰性/PDH陽性MDVsを示す.mito-DsRed陽性のMDVsは観察されなかった.
DRP1(DNM1L, DLP1)は細胞質に存在するGTPaseである19, 20).ミトコンドリアを囲むように多量体を形成し,GTPase活性依存的にミトコンドリアの切断を促進する.DRP1の遺伝子発現抑制やGTPaseのドミナントネガティブ体を発現させると,ミトコンドリアの分裂が抑制され,長く伸長したミトコンドリアネットワークが形成される(図2).このような条件下でもMDVsの数は影響されないことから,MDVsの放出はDRP1非依存的であると考えられていた5, 6).しかし,McBride博士自身らのグループからDRP1依存的なMDVsの放出が報告された16).これまでのDRP1抑制系とは異なり,CRISPR/Cas9システムでノックアウトしたところ,ある種のMDVsの形成・放出が有意に抑制された.このときのDRP1のミトコンドリアへの集積には,ミトコンドリア分裂と同様の受容体が関与していた.興味深いことに,DRP1依存的なMDVsの放出は突起のように伸長したミトコンドリアの先端で行われており,その特徴はperipheral fissionと類似している18).ミトコンドリア本体から出るMDVsも観察されているので,カーゴや細胞種などにより分子機構が異なり,DRP1依存的・非依存的な両方のMDVs制御機構が存在することが示唆されている.また,DRP1の発現抑制やドミナントネガティブ体の発現は,MDVsと断片化したミトコンドリアを区別するためには依然として有効な手段ではあるので,MDVsの予備的な検討としては推奨される実験条件である.
その他のMDVsの特徴を以下に記す.
ミトコンドリアは脂質組成の異なる内膜・外膜の二重膜から形成され,内膜はクリステ構造と呼ばれるひだ状の構造を発達させている.また,内膜の物質透過性は外膜と比べ低く,電子伝達系によって膜間部に汲み出されたプロトン濃度勾配により膜電位を形成している.透過型電子顕微鏡による微細構造の解析から,ペルオキシソーム・リソソーム経路ともに外膜のみの一重膜,外膜・内膜を含んだ二重膜のMDVsが観察されている5, 6).
ミトコンドリアは核とは独立した独自のDNAを有している.近年,細胞質に存在するmtDNAが炎症系のシグナルを活性化することが明らかになってきた21–23).その一部の機構としてフマル酸の蓄積により誘導されるMDVsを介したmtDNAの放出が見いだされ,細胞内代謝と免疫シグナリングをつなぐMDVsの新たな役割が明らかとなった24, 25).一方で,定常状態の培養細胞やウシ心臓より精製したミトコンドリアを用いたin vitro再構成系の実験では確認されなかったことから,シグナルを受容しカーゴ選択的にmtDNAが放出されていると考えられる26, 27).
MDVsの研究が進まなかった理由の一つとして,これまで検出されていなかったことが考えられる.免疫蛍光染色などによりミトコンドリアの形態を観察するとき,細胞固定前のリン酸緩衝液(PBS)での洗浄はミトコンドリアの形態に影響を及ぼすが,MDVsは検出自体されなくなってしまう.MDVs検出のためには,培養培地を取り除いた直後に37°Cに温めておいた5%パラホルムアルデヒド(PFA)溶液を加え,37°Cで15分間固定する.マルチウェルプレートで行う場合は,1ウェルずつ培地とPFAの交換を行うことを推奨したい.PFAは20%で調製後,−20°Cで保存し固定前に融解しPBSで希釈して使用する.凍結融解は繰り返さない.メタノールやアセトンによる脱水固定でもMDVsは保持されない.
以上がMDVsの一般的な特徴と検出のためのヒントである.本稿ではMDVsの輸送先ごとに概説していき,最後にMDVsの進化的な側面について述べる.
McBride博士らの研究グループはMDVsの誘導条件を探索する過程でミトコンドリア呼吸能阻害などの酸化ストレス下でMDVsの形成が促され,それらのMDVsはリソソームへと輸送されることを見いだした6).さらに,単離精製したミトコンドリアを用いたin vitro再構築系により,MDVsには酸化されたタンパク質が多く含まれていることが明らかとなり,酸化タンパク質(おそらく脂質も)を選択的に除去する品質管理機構の一つであることが示された26).活性酸素種の産生などによる心筋毒性が知られているドキソルビシンで処理したマウス心臓の電子顕微鏡像においてもMDVsの形成が観察され,生体内においても酸化ストレスに対するミトコンドリア品質管理機構としてMDVsが働いていることが支持されている28).ミトコンドリアはマトリックスや内膜におけるプロテアーゼ,外膜におけるユビキチン・プロテアソーム系などによるタンパク質レベルから,ミトコンドリア特異的オートファジーであるマイトファジーのようにオルガネラレベルの品質管理機構により恒常性が保たれている.ATG5やLC3といったオートファジー制御因子とも独立的に機能していることから,MDVsはマイトファジーとは異なる新たな膜動態を介したミトコンドリア品質管理機構である6).また,マイトファジー阻害下においては,MDVsが代償的にミトコンドリア品質管理を担っていることも観察されており,ミトコンドリア機能維持において重要な機構の一つであるといえる29).若年性パーキンソン病の原因遺伝子であるPARK2(Parkin)とPARK6(PINK1)はある種のマイトファジーにおいて中心的な役割を果たしているが,酸化ストレス誘導性のMDVs生成も正に制御している30).局所的あるいは部分的な除去はMDVs,オルガネラごとの除去はマイトファジーが担っていることが予想されるが,傷害やストレスレベルの感知機構や両経路の使い分けなどについては未解明な部分が多く残されている.
上述のようにPINK1/Parkinは傷害ミトコンドリアを丸ごと分解するマイトファジーの主要経路であるので,PINK1/Parkinの病的バリアントによるミトコンドリア品質管理機構の破綻がパーキンソン病の原因の一つであるとされていた.PINK1/Parkin制御によるMDVsとパーキンソン病の関連については品質管理とは異なる説が提唱されているので,ここで紹介したい(図3).
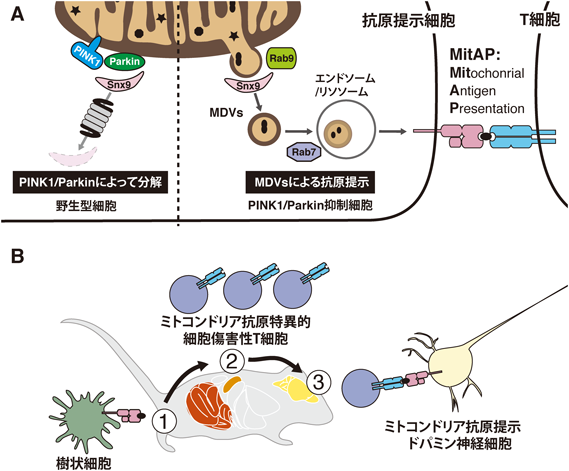
(A) MitAPの分子機構.野生型細胞(左)ではSnx9の分解がPINK1/Parkinによって促進されているため,MitAPが抑制されている.PINK1/Parkin抑制下(右)ではSnx9によるミトコンドリア抗原を含んだMDVsの放出が促され,Rab9・Rab7を介したMitAPが過度に促進される.(B) PINK1ノックアウトマウスのパーキンソン病発症モデル.腸内感染により,樹状細胞によるミトコンドリア抗原提示が誘導される(①).脾臓において,ミトコンドリア抗原を認識する細胞傷害性T細胞が増殖する(②).細胞傷害性T細胞が中枢神経系へ浸潤し,ミトコンドリア抗原を提示しているドパミン神経細胞を攻撃し,神経細胞死を誘導する(③).
カナダ・モントリオール大学のDesjardins博士らの研究グループは細胞内局在特異的な抗原提示経路を調べるために,単純ヘルペスウイルスエンベロープの糖タンパク質B(gB)をミトコンドリアマトリックス(mito-gB),核膜(NE-gB),細胞質(Cyto-gB)に発現させたマクロファージ系の細胞を用いて抗原提示経路について解析したところ,mito-gBの抗原提示が特異的に熱ストレスにより誘導された8).このときの抗原提示経路はマイトファジーおよびオートファジーと独立した経路であり,mito-gBに加え,内在性のミトコンドリアマトリックスタンパク質であるOGDH(α-ケトグルタル酸デヒドロゲナーゼ)もカーゴ選択的にMDVsとして放出され,細胞表面に提示される.カーゴの選択的取り込み機構の詳細は不明であるが,ミトコンドリアからの放出はSnx9やRab9,後期エンドソーム/リソソームへの輸送はRab7といったエンドサイトーシスなどにも関わる分子が制御している.このようなMDVs依存的なミトコンドリア特異的な抗原提示経路はMitAP(mitochondrial antigen presentation)と命名された(図3A)8).リソソームへのMDVs経路ということでPINK1/Parkinによる活性化が予想されたが,正常な細胞ではSnx9を分解することによりMitAPを抑制的に制御していることが明らかとなった(図3A左).すなわち,病原性変異によるPINK1/Parkinの機能低下が過剰なMitAP,炎症反応を誘導している可能性が示唆された(図3A右).MitAPとパーキンソン病の関係を調べるために,PINK1ノックアウトマウスにげっ歯類の感染性大腸炎モデルとしてよく用いられるグラム陰性細菌であるCitrobacter rodentiumを感染させたところ,運動能力の低下やドーパミン作動性神経細胞軸索上の結節状構造異常などのパーキンソン病様の症状を呈した9).腸内感染により樹状細胞におけるOGDH抗原提示が促進され,それを認識する細胞傷害性T細胞が増殖する.中枢神経系ではドパミン神経がOGDH抗原を提示しており,中枢神経系へ浸潤してきた細胞傷害性T細胞による攻撃によってドーパミン神経細胞死が誘導されていることが示された(図3B).
これまでの研究ではPINK1やParkinを純粋にノックアウトしただけではパーキンソン病様の表現型を呈さないことが,パーキンソン病モデルマウスの一つの課題であった31).我々“野生”のヒトは常に環境中の感染源にさらされているが,過度な炎症は起こらない程度に免疫系が働き,感染症から守られている.一方で多くの場合,実験動物はC. rodentium等も含めた感染源が管理されたSPF(specific pathogen free,特定病原体非存在)下で飼育されている.パーキンソン病モデルマウスの生育環境はクリーンすぎるために,パーキンソン病を発症しづらかったのかもしれない.上記の結果よりパーキンソン病はParkinやPINK1など原因遺伝子の病的バリアントによる機能低下に加え,腸内感染等による炎症が契機となり,自己免疫疾患様の機構で発症する新たなモデルが提唱された.しかし,実際に個体における腸内感染によるドパミン神経細胞死は確認できていないことから,今後さらなる解析と詳細な発症分子機序の解明が期待される.
MDVsの最初の発見はペルオキシソームへの輸送であった.MAPL(MUL1, MULAN)はミトコンドリア外膜とペルオキシソーム膜に局在するE3ユビキチンリガーゼである32).基質のSUMO化やユビキチン化を行うことにより,ミトコンドリアの形態やアポトーシス,マイトファジー,胆汁酸輸送を制御している32–37).McBride博士らのグループはMAPL過剰発現細胞のミトコンドリア形態を透過型電子顕微鏡により解析したところ,ミトコンドリアとつながった,あるいは近接した特徴的な小胞様の構造を発見した5).このときの細胞を異なる複数のミトコンドリアマーカーで多重標識すると,MAPL陽性の構造は他のマーカーを含まなかった.また,ミトコンドリア分裂因子であるDRP1抑制下でも同様の構造が観察されることから,MAPLがカーゴ選択的に小胞に取り込まれて,ミトコンドリア分裂とは異なる機構で放出されることが示された.そこで,非対称分裂したミトコンドリアの断片と区別しMDVsと名づけられた.その後,MAPLの結合タンパク質の網羅的な解析により,レトロマータンパク質の一つであるVPS35が同定された38).VPS35の遺伝子発現を抑制すると,MAPL陽性のペルオキシソーム数が減少するので,少なくとも一部のMAPLはMDVsによってミトコンドリアからペルオキシソームに運ばれている.しかし,MAPLがミトコンドリアを経由してペルオキシソームに輸送されることの必要性など,ペルオキシソームへのMDVs輸送の生理的意義については多くの謎に包まれたままである.現在までに既存のペルオキシソームへと運ばれるMDVsのカーゴはMAPLしか同定されておらず,今後プロテオームなどの網羅的な解析によりカーゴや制御因子が同定されることが期待される.
ペルオキシソームは細胞内外の環境に応じて数や形態を変化させるが,細胞内の数は成長・分裂サイクルによる増殖と分解のバランスにより保たれている39, 40).また.実験的な条件においては,他の膜構造から前駆小胞が形成されde novo合成されるという特徴を有する.de novo合成の解析は出芽酵母Saccharomyces cerevisiaeのペルオキシソーム欠損変異株を使った研究が先行して行われ,小胞体がペルオキシソーム膜の起源であるモデルが確立されつつある41, 42).一方,Hansenula polymorphaのペルオキシソーム欠損細胞株ではペルオキシソーム前駆体となる膜構造が存在し,小胞体を介さずにde novo合成が行われることが報告されており,de novo合成における膜の起源の多様性も示唆されている43).哺乳類細胞では,一部のペルオキシソーム膜タンパク質が小胞体を経由してペルオキシソームへ輸送されている報告もあり,哺乳類においても小胞体起源説が広く受け入れられている44, 45).しかし,哺乳類ペルオキシソーム欠損培養細胞では複数のペルオキシソーム膜タンパク質がミトコンドリアに局在することが報告されているにもかかわらず,それらはアーティファクトとして解釈されていた46, 47).筆者らの研究グループはZellweger症候群由来ペルオキシソーム欠損線維芽細胞を用いて,de novo合成の過程を段階的に解析した.PEX3変異により,PEX3の発現およびペルオキシソームを欠失している細胞にPEX3を発現させた細胞を透過型電子顕微鏡で観察するとミトコンドリアに小胞様の構造が観察された.さらに,金コロイドを用いた免疫電子顕微鏡法を行ったところ,PEX3が小胞様の構造に蓄積していることも確認された.ライブセルイメージングにより,PEX3がミトコンドリアの一部に集積し,放出されることが観察され,ミトコンドリアからペルオキシソーム前駆小胞が形成されていることが示された.また,PEX3発現誘導後の経時的なサンプリングによる成熟化過程の解析により小胞体由来の前駆小胞との融合が必要であることも明らかになり,ペルオキシソームde novo合成の新たな仮説であるハイブリッドモデルが提唱された(図4)48).同時にペルオキシソーム前駆小胞という新たなMDVsの役割が示された.しかし,分子制御機構に加え,最も重要な“生理的にde novo合成は起きているか?”という問いにはいまだに答えは出ておらず,大きな課題が残されている.
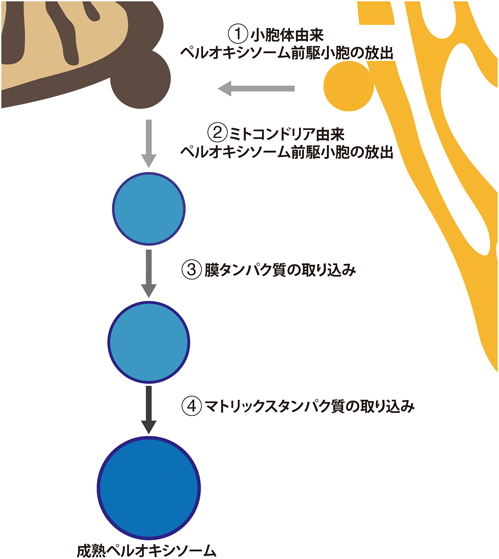
①小胞体よりペルオキシソーム前駆小胞が放出される.②ミトコンドリア由来ペルオキシソーム前駆小胞が放出される.③④細胞質で合成された膜タンパク質やマトリックスタンパク質を取り込みながら成熟ペルオキシソームとなる.
ペルオキシソームとミトコンドリアは脂肪酸酸化や活性酸素種の除去などの機能に加え,形態制御,オートファジー,自然免疫シグナル伝達,転写因子など多くの分子機構も共有しているが,その相互制御や意義についての十分な理解は得られていない49).既存のペルオキシソームへの輸送,de novo合成経路におけるMDVsのカーゴや誘導条件の同定などができれば,両オルガネラの新たな関係性も明らかになるかもしれない.
MDVsは不要なものをミトコンドリアから除去するだけでなく,必要なものもミトコンドリアから輸送する.以下にリソソームやペルオキシソーム以外の輸送先で,MDVsの機能や生理的意義の一端が示された報告例を記す.
過度な活性酸素種は酸化ストレスを引き起こすが,活性酸素種はシグナル伝達や細胞内の感染源に対する殺菌作用を有し,細胞内においては必要な分子である.多剤耐性であるメチシリン耐性黄色ブドウ球菌(methicillin-resistant Staphylococcus Aureus:MRSA)はさまざまな感染症の原因となる.生体内ではマクロファージに貪食された後に,ファゴソームに取り込まれ,殺菌・分解される.感染により小胞体ストレスが誘導され,ミトコンドリアにおける活性酸素種の産生も促されるが,どのようにミトコンドリア由来の活性酸素種がファゴソーム内に運ばれるかはわかっていなかった.MRSA感染マクロファージにおいて小胞体ストレスの下流で過酸化水素を含んだMDVsの形成・放出が促され,ファゴソームへ輸送されることが見いだされた50).このときにミトコンドリアマトリックスタンパク質であるSOD2も同時に輸送され,ファゴソーム内におけるさらなる過酸化水素の産生を促し,MRSAを除去する.また,これらのMDVs経路はPINK1/Parkinによって正に制御されているが,パーキンソン病との関連は明らかとなっていない.
骨芽細胞の分化・成熟過程において,ミトコンドリアは伸長,ドーナッツ様の形態,分裂とダイナミックに形態を変化させる.分裂とともに呼吸能も低下していき,最終的にはマイトファジーによって分解される.成熟骨芽細胞から分裂とともにMDVsの形成・放出が促され,CD38/cADPR(cluster of differentiation 38/cyclic adenosine diphosphate-ribose)調節性の細胞外小胞によって細胞外に放出される.培養液中のMDVsを精製し,培養骨芽細胞に添加すると骨への分化が促されることから,MDVsが骨芽細胞の成熟化に関与していることが明らかとなった51).外膜のみの一重膜,外膜・内膜からなる二重膜のMDVsで同様の分化促進作用が認められたが,分化誘導因子となるようなカーゴについては同定されていない.
このようにMDVsが細胞機能の維持や分化に積極的に関与していることも近年徐々に明らかになってきている.また,細胞外小胞にミトコンドリアあるいは一部のミトコンドリアタンパク質が取り込まれていることは以前より知られていたが,一連の輸送にMDVsが関与していることも示唆されている.発見当初は全体のごく一部,ミトコンドリアの特殊な現象だと考えられていたが細胞内膜輸送の主要経路の一つであることが徐々に明らかになりつつある.今後のMDVsのさらなる生理的な機能の解明が期待される.
MDVsの発見当時は実験の人工産物であるとの見解が多勢を占めていた.特に,ミトコンドリア動態を専門としていた研究者の方々は「見たことがない」というのが端的な理由であったと予想される.ミトコンドリアの進化的な祖先であると考えられているグラム陰性細菌であるアルファプロテオバクテリオは,恒常的に小胞[細胞外小胞(extracellular vesicles)]を放出している14).この観点から,MDVsは実は進化的にも保存されたミトコンドリアの膜動態の一つであるとも考えられる.ミトコンドリアと細胞内膜構造の発達・進化の関係性は活発に議論されているが,宿主が貪食によりアルファプロテオバクテリアを取り込んだ,すなわちミトコンドリア共生以前にある程度の細胞内膜構造が発達したとする説と,ミトコンドリアの共生後に細胞内膜構造が発達したとする説が相克している52).後者の説に関して,宿主内で放出されたMDVsが細胞内膜構造の発達に寄与したとの説が提唱された(図5)53).この説は細胞膜の脂質組成が古細菌ではなく,細菌に類似していることも説明する.また,それを裏づけるように,真核生物との祖先に最も近縁と考えられる古細菌MK-D1が単離培養された54).MK-D1は真核生物固有と考えられていたアクチンやESCRTIIIなどの遺伝子を多数持つが,重要なことに目立った細胞内膜構造を持っていなかった.また,独特な突起を介し周りの細菌と代謝物の輸送などの相互作用をすることにより,共生し生存している可能性が示された.MK-D1は突起を用いて,ミトコンドリアの祖先となる細菌を細胞内に取り込み進化した,とされる真核生物説の進化に関する新たな説,“Entangle-Engulf-Endogenize(E3)model”が提唱された.この説もミトコンドリアが取り込まれた後に,細胞内膜構造が発達していった説を支持する.一方で,ペルオキシソームの進化的な起源についての知見は少なく,多くの謎に包まれたままである.独立したグループのプロテオームの結果より,ミトコンドリアとペルオキシソームタンパク質の進化的起源が類似していることが示されている55, 56).上記のモデルにより提唱された細胞内膜構造の発達にはペルオキシソームも含まれている可能性も十分にあり,筆者らが発見した前述のde novo合成の過程はこのような進化の名残かもしれない.進化の過程を実験的に再現・証明することは難しいが,さまざまな生物種のMDVsとオルガネラの関係を調べることにより,細胞内膜構造の発達の謎に迫ることが期待される.
(A)宿主となる古細菌(水色)が突起を介して,ミトコンドリア祖先細菌(茶色)と相互作用し,栄養物などの輸送や交換を行う.(B)宿主に取り込まれた後も,ミトコンドリア祖先細菌は細胞外小胞の放出を維持している.(C)細胞膜の組成が細菌由来に置き換わり,核膜も形成される.(D)小胞体やリソソーム,ペルオキシソームといった他のオルガネラも形成される.文献53, 54のモデルを参考に作成した.
MDVsが初めて報告されてから,約15年が経つ.発見後しばらくはMcBride博士らの研究グループの独壇場であったが,ここ数年ようやくそれ以外のグループより報告されるようになった.上記のように,目的地,カーゴ,制御機構などさまざまな特徴を持ったMDVsが次々と発見されている.哺乳類での解析を中心に疾患との関わりも徐々に明らかになってきた.植物での報告もあるように,さまざまな生物で広く保存されている可能性もあり,今後はさまざまな生物種におけるMDVsの研究も期待される57).MDVsの実体を明らかにすることにより,ミトコンドリアの新たな側面や生命の基盤の理解につながるかもしれない.今後は,特に若手研究者の方々に参入,挑戦していただきその理解を深めていきたい.
本総説は2023年生化学会奨励賞受賞に際し,寄稿させていただいた.これまで研究をご指導してくださいました先生方,共に研鑽してきた友人,よくわからないことを理解し支えてくれている家族には心より感謝申し上げたい.
本総説は2023年度奨励賞を受賞した.
1) Suomalainen, A. & Battersby, B.J. (2018) Mitochondrial diseases: The contribution of organelle stress responses to pathology. Nat. Rev. Mol. Cell Biol., 19, 77–92.
2) Murayama, K., Shimura, M., Liu, Z., Okazaki, Y., & Ohtake, A. (2019) Recent topics: The diagnosis, molecular genesis, and treatment of mitochondrial diseases. J. Hum. Genet., 64, 113–125.
3) Giacomello, M., Pyakurel, A., Glytsou, C., & Scorrano, L. (2020) The cell biology of mitochondrial membrane dynamics. Nat. Rev. Mol. Cell Biol., 21, 204–224.
4) Borcherding, N. & Brestoff, J.R. (2023) The power and potential of mitochondria transfer. Nature, 623, 283–291.
5) Neuspiel, M., Schauss, A.C., Braschi, E., Zunino, R., Rippstein, P., Rachubinski, R.A., Andrade-Navarro, M.A., & McBride, H.M. (2008) Cargo-selected transport from the mitochondria to peroxisomes is mediated by vesicular carriers. Curr. Biol., 18, 102–108.
6) Soubannier, V., McLelland, G.-L., Zunino, R., Braschi, E., Rippstein, P., Fon, E.A., & McBride, H.M. (2012) A vesicular transport pathway shuttles cargo from mitochondria to lysosomes. Curr. Biol., 22, 135–141.
7) Ryan, T.A., Phillips, E.O., Collier, C.L., Jb Robinson, A., Routledge, D., Wood, R.E., Assar, E.A., & Tumbarello, D.A. (2020) Tollip coordinates Parkin-dependent trafficking of mitochondrial-derived vesicles. EMBO J., 39, e102539.
8) Matheoud, D., Sugiura, A., Bellemare-Pelletier, A., Laplante, A., Rondeau, C., Chemali, M., Fazel, A., Bergeron, J.J., Trudeau, L.E., Burelle, Y., et al. (2016) Parkinson’s disease-related proteins PINK1 and Parkin repress mitochondrial antigen presentation. Cell, 166, 314–327.
9) Matheoud, D., Cannon, T., Voisin, A., Penttinen, A.-M., Ramet, L., Fahmy, A.M., Ducrot, C., Laplante, A., Bourque, M.J., Zhu, L., et al. (2019) Intestinal infection triggers Parkinson’s disease-like symptoms in Pink1−/− mice. Nature, 571, 565–569.
10) Li, B., Zhao, H., Wu, Y., Zhu, Y., Zhang, J., Yang, G., Yan, Q., Li, J., Li, T., & Liu, L. (2020) Mitochondrial-derived vesicles protect cardiomyocytes against hypoxic damage. Front. Cell Dev. Biol., 8, 214.
11) Crewe, C., Funcke, J.B., Li, S., Joffin, N., Gliniak, C.M., Ghaben, A.L., An, Y.A., Sadek, H.A., Gordillo, R., Akgul, Y., et al. (2021) Extracellular vesicle-based interorgan transport of mitochondria from energetically stressed adipocytes. Cell Metab., 33, 1853–68.e11.
12) Mondet, J., Lo Presti, C., Chevalier, S., Bertrand, A., Tondeur, S., Blanchet, S., Mc Leer, A., Pernet-Gallay, K., & Mossuz, P. (2021) Mitochondria in human acute myeloid leukemia cell lines have ultrastructural alterations linked to deregulation of their respiratory profiles. Exp. Hematol., 98, 53–62.e3.
13) Pouikli, A., Parekh, S., Maleszewska, M., Nikopoulou, C., Baghdadi, M., Tripodi, I., Folz-Donahue, K., Hinze, Y., Mesaros, A., Hoey, D., et al. (2021) Chromatin remodeling due to degradation of citrate carrier impairs osteogenesis of aged mesenchymal stem cells. Nat. Aging, 1, 810–825.
14) Kaplan, M., Chreifi, G., Metskas, L.A., Liedtke, J., Wood, C.R., Oikonomou, C.M., Nicolas, W.J., Subramanian, P., Zacharoff, L.A., Wang, Y., et al. (2021) In situ imaging of bacterial outer membrane projections and associated protein complexes using electron cryo-tomography. eLife, 10, e73099.
15) Sugiura, A., McLelland, G.-L., Fon, E.A., & McBride, H.M. (2014) A new pathway for mitochondrial quality control: Mitochondrial-derived vesicles. EMBO J., 33, 1–15.
16) König, T., Nolte, H., Aaltonen, M.J., Tatsuta, T., Krols, M., Stroh, T., Langer, T., & McBride, H.M. (2021) MIROs and DRP1 drive mitochondrial-derived vesicle biogenesis and promote quality control. Nat. Cell Biol., 23, 1271–1286.
17) Twig, G., Elorza, A., Molina, A.J., Mohamed, H., Wikstrom, J.D., Walzer, G., Stiles, L., Haigh, S.E., Katz, S., Las, G., et al. (2008) Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J., 27, 433–446.
18) Kleele, T., Rey, T., Winter, J., Zaganelli, S., Mahecic, D., Perreten Lambert, H., Ruberto, F.P., Nemir, M., Wai, T., Pedrazzini, T., et al. (2021) Distinct fission signatures predict mitochondrial degradation or biogenesis. Nature, 593, 435–439.
19) Shin, H.W., Shinotsuka, C., Torii, S., Murakami, K., & Nakayama, K. (1997) Identification and subcellular localization of a novel mammalian dynamin-related protein homologous to yeast Vps1p and Dnm1p. J. Biochem., 122, 525–530.
20) Smirnova, E., Shurland, D.L., Ryazantsev, S.N., & van der Bliek, A.M. (1998) A human dynamin-related protein controls the distribution of mitochondria. J. Cell Biol., 143, 351–358.
21) West, A.P., Khoury-Hanold, W., Staron, M., Tal, M.C., Pineda, C.M., Lang, S.M., Bestwick, M., Duguay, B.A., Raimundo, N., MacDuff, D.A., et al. (2015) Mitochondrial DNA stress primes the antiviral innate immune response. Nature, 520, 553–557.
22) White, M.J., McArthur, K., Metcalf, D., Lane, R.M., Cambier, J.C., Herold, M.J., van Delft, M.F., Bedoui, S., Lessene, G., Ritchie, M.E., et al. (2014) Apoptotic caspases suppress mtDNA-induced STING-mediated type I IFN production. Cell, 159, 1549–1562.
23) Rongvaux, A., Jackson, R., Harman, C.C., Li, T., West, A.P., de Zoete, M.R., Wu, Y., Yordy, B., Lakhani, S.A., Kuan, C.Y., et al. (2014) Apoptotic caspases prevent the induction of type I interferons by mitochondrial DNA. Cell, 159, 1563–1577.
24) Hooftman, A., Peace, C.G., Ryan, D.G., Day, E.A., Yang, M., McGettrick, A.F., Yin, M., Montano, E.N., Huo, L., Toller-Kawahisa, J.E., et al. (2023) Macrophage fumarate hydratase restrains mtRNA-mediated interferon production. Nature, 615, 490–498.
25) Zecchini, V., Paupe, V., Herranz-Montoya, I., Janssen, J., Wortel, I.M.N., Morris, J.L., Ferguson, A., Chowdury, S.R., Segarra-Mondejar, M., Costa, A.S.H., et al. (2023) Fumarate induces vesicular release of mtDNA to drive innate immunity. Nature, 615, 499–506.
26) Soubannier, V., Rippstein, P., Kaufman, B.A., Shoubridge, E.A., & McBride, H.M. (2012) Reconstitution of mitochondria derived vesicle formation demonstrates selective enrichment of oxidized cargo. PLoS One, 7, e52830.
27) Sánchez, M.I., Vida, Y., Pérez-Inestrosa, E., Mascareñas, J.L., Vázquez, M.E., Sugiura, A., & Martínez-Costas, J. (2020) MitoBlue as a tool to analyze the mitochondria-lysosome communication. Sci. Rep., 10, 3528.
28) Cadete, V.J.J., Deschenes, S., Cuillerier, A., Brisebois, F., Sugiura, A., Vincent, A., Turnbull, D., Picard, M., McBride, H.M., & Burelle, Y. (2016) Formation of mitochondrial-derived vesicles is an active and physiologically relevant mitochondrial quality control process in the cardiac system. J. Physiol., 594, 5343–5362.
29) Towers, C.G., Wodetzki, D.K., Thorburn, J., Smith, K.R., Caino, M.C., & Thorburn, A. (2021) Mitochondrial-derived vesicles compensate for loss of LC3-mediated mitophagy. Dev. Cell, 56, 2029–2042.e5.
30) McLelland, G.-L., Soubannier, V., Chen, C.X., McBride, H.M., & Fon, E.A. (2014) Parkin and PINK1 function in a vesicular trafficking pathway regulating mitochondrial quality control. EMBO J., 33, 282–295.
31) Paul, S. & Pickrell, A.M. (2021) Hidden phenotypes of PINK1/Parkin knockout mice. Biochim. Biophys. Acta, Gen. Subj., 1865, 129871.
32) Braschi, E., Zunino, R., & McBride, H.M. (2009) MAPL is a new mitochondrial SUMO E3 ligase that regulates mitochondrial fission. EMBO Rep., 10, 748–754.
33) Li, W., Bengtson, M.H., Ulbrich, A., Matsuda, A., Reddy, V.A., Orth, A., Chanda, S.K., Batalov, S., & Joazeiro, C.A. (2008) Genome-wide and functional annotation of human E3 ubiquitin ligases identifies MULAN, a mitochondrial E3 that regulates the organelle’s dynamics and signaling. PLoS One, 3, e1487.
34) Prudent, J., Zunino, R., Mattie, S., Gordon, C., McBride, H.M., Sugiura, A., et al. (2015) MAPL SUMOylation of Drp1 stabilizes an ER/mitochondrial platform required for cell death article MAPL SUMOylation of Drp1 stabilizes an ER/mitochondrial platform required for cell death. Mol. Cell, 59, 941–955.
35) Goyon, V., Besse-Patin, A., Zunino, R., Ignatenko, O., Nguyen, M., Coyaud, E., Lee, J.M., Nguyen, B.N., Raught, B., & McBride, H.M. (2023) MAPL loss dysregulates bile and liver metabolism in mice. EMBO Rep., 24, e57972.
36) Ambivero, C.T., Cilenti, L., Main, S., & Zervos, A.S. (2014) Mulan E3 ubiquitin ligase interacts with multiple E2 conjugating enzymes and participates in mitophagy by recruiting GABARAP. Cell. Signal., 26, 2921–2929.
37) Li, J., Qi, W., Chen, G., Feng, D., Liu, J., Ma, B., Zhou, C., Mu, C., Zhang, W., Chen, Q., et al. (2015) Mitochondrial outer-membrane E3 ligase MUL1 ubiquitinates ULK1 and regulates selenite-induced mitophagy. Autophagy, 11, 1216–1229.
38) Braschi, E., Goyon, V., Zunino, R., Mohanty, A., Xu, L., & McBride, H.M. (2010) Vps35 mediates vesicle transport between the mitochondria and peroxisomes. Curr. Biol., 20, 1310–1315.
39) Lazarow, P.B. & Fujiki, Y. (1985) Biogenesis of peroxisomes. Annu. Rev. Cell Biol., 1, 489–530.
40) Yagita, Y., Abe, Y., & Fujiki, Y. (2022) De novo formation and maintenance of mammalian peroxisomes in cultured PEX16-knockout cells generated by CRISPR/Cas9. J. Cell Sci., 135, jcs258377.
41) Hoepfner, D., Schildknegt, D., Braakman, I., Philippsen, P., & Tabak, H.F. (2005) Contribution of the endoplasmic reticulum to peroxisome formation. Cell, 122, 85–95.
42) van der Zand, A., Gent, J., Braakman, I., & Tabak H.F. (2012) Biochemically distinct vesicles from the endoplasmic reticulum fuse to form peroxisomes. Cell, 149, 397–409.
43) Knoops, K., Manivannan, S., Cepińska, M.N., Krikken, A.M., Kram, A.M., Veenhuis, M., & van der Klei, I.J. (2014) Preperoxisomal vesicles can form in the absence of Pex3. J. Cell Biol., 204, 659–668.
44) Aranovich, A., Hua, R., Rutenberg, A.D., & Kim, P.K. (2014) PEX16 contributes to peroxisome maintenance by constantly trafficking PEX3 via the ER. J. Cell Sci., 127, 3675–3686.
45) Tabak, H.F., Murk, J.L., Braakman, I., & Geuze, H.J. (2003) Peroxisomes start their life in the endoplasmic reticulum. Traffic, 4, 512–518.
46) Sacksteder, K.A., Jones, J.M., South, S.T., Li, X., Liu, Y., & Gould, S.J. (2000) PEX19 binds multiple peroxisomal membrane proteins, is predominantly cytoplasmic, and is required for peroxisome membrane synthesis. J. Cell Biol., 148, 931–944.
47) Toro, A.A., Araya, C.A., Córdova, G.J., Arredondo, C.A., Cárdenas, H.G., Moreno, R.E., Venegas, A., Koenig, C.S., Cancino, J., Gonzalez, A., et al. (2009) Pex3p-dependent peroxisomal biogenesis initiates in the endoplasmic reticulum of human fibroblasts. J. Cell. Biochem., 107, 1083–1096.
48) Sugiura, A., Mattie, S., Prudent, J., & McBride, H.M. (2017) Newly born peroxisomes are a hybrid of mitochondrial and ER-derived pre-peroxisomes. Nature, 542, 251–254.
49) Schrader, M., Costello, J., Godinho, L.F., & Islinger, M. (2015) Peroxisome-mitochondria interplay and disease. J. Inherit. Metab. Dis., 38, 681–702.
50) Abuaita, B.H., Schultz, T.L., & O’Riordan, M.X. (2018) Mitochondria-derived vesicles deliver antimicrobial reactive oxygen species to control phagosome-localized staphylococcus aureus. Cell Host Microbe, 24, 625–636.
51) Suh, J., Kim, N.K., Shim, W., Lee, S.H., Kim, H.J., Moon, E., Sesaki, H., Jang, J.H., Kim, J.E., & Lee, Y.S. (2023) Mitochondrial fragmentation and donut formation enhance mitochondrial secretion to promote osteogenesis. Cell Metab., 35, 345–360.e7.
52) Martin, W.F., Tielens, A.G.M., Mentel, M., Garg, S.G., & Gould, S.B. (2017) The physiology of phagocytosis in the context of mitochondrial origin. Microbiol. Mol. Biol. Rev., 81, e00008–e00017.
53) Gould, S.B., Garg, S.G., & Martin, W.F. (2016) Bacterial vesicle secretion and the evolutionary origin of the eukaryotic endomembrane system. Trends Microbiol., 24, 525–534.
54) Imachi, H., Nobu, M.K., Nakahara, N., Morono, Y., Ogawara, M., Takaki, Y., Takano, Y., Uematsu, K., Ikuta, T., Ito, M., et al. (2020) Isolation of an archaeon at the prokaryote-eukaryote interface. Nature, 577, 519–525.
55) Szklarczyk, R. & Huynen, M.A. (2010) Mosaic origin of the mitochondrial proteome. Proteomics, 10, 4012–4024.
56) Hartwig, S., Knebel, B., Goeddeke, S., Koellmer, C., Jacob, S., Nitzgen, U., Passlack, W., Schiller, M., Dicken, H.D., Haas, J., et al. (2013) So close and yet so far: Mitochondria and peroxisomes are one but with specific talents. Arch. Physiol. Biochem., 119, 126–135.
57) Yamashita, A., Fujimoto, M., Katayama, K., Yamaoka, S., Tsutsumi, N., & Arimura, S. (2016) Formation of Mitochondrial Outer Membrane Derived Protrusions and Vesicles in Arabidopsis thaliana. PloS one, 11(1), e0146717.

順天堂大学大学院医学研究科 講師.博士(生命科学).
1984年神奈川県に生る.2007年東京薬科大学生命科学部卒業.12年同大学院生命科学研究科修了.12~16カナダ・マギル大学研究員.14~16日本学術振興会海外特別研究員(兼任).16~17年東京薬科大学プロジェクト研究員.17~20年神戸大学特命助教.20年より現職.
研究テーマと抱負ペルオキシソームの出生が知りたいです.
趣味裁縫.

This page was created on 2024-03-21T19:16:49.195+09:00
This page was last modified on 2024-04-17T08:32:39.000+09:00
このサイトは(株)国際文献社によって運用されています。