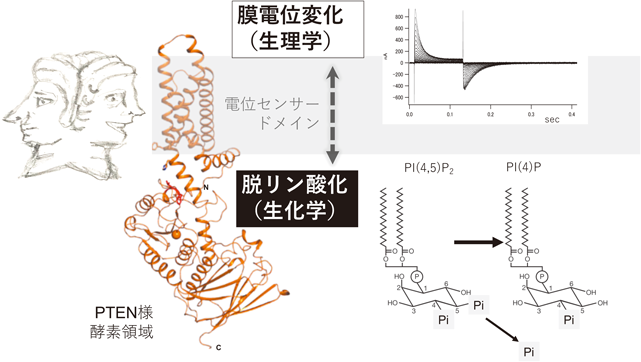
ヤリイカの神経軸索に始まるイオンチャネル仮説とその後の分子の発見や,神経伝達物質の放出に関する量子仮説とその後の分子機構の解明など,これまで,計測技術の進歩により生命現象が詳しく解明され,背景にある原理と分子実体が明らかにされてきた.本稿で述べるVSPの研究は,こうした正統的な流れと大きく異なっている.VSPは,約20年前に海産動物ホヤのゲノム解析を契機にアノテーションの過程で,電気信号と化学信号を共役させるユニークな分子として偶然に見いだされた1)(図1).これまでの研究の意義と現状をまとめる.
被嚢動物であるホヤは脊椎動物に最も近縁である無脊椎動物であり,長く初期発生の研究の対象となってきた.ショウジョウバエの遺伝学を確立したMorganもホヤの研究を行っていたことがある.2002年にカタユウレイボヤの全ゲノムの塩基配列が明らかにされ2),そのころイオンチャネル関連遺伝子のアノテーションを行う機会3)に恵まれた.その際,既存のどのイオンチャネル分子のカテゴリーにも属さない複数のイオンチャネル関連遺伝子に遭遇した.その一つはN末端側が電位依存性Na+チャネルと相同性を示す(図2)一方で,C末端側はPTENとの相同性を示していた.この分子はすでにヒトや齧歯類から膜貫通領域を持つPTEN様分子として発見され,機能未知の酵素(TPTE, TPIP, PTEN2などの名称)としていくつかの論文が出されていたが,膜貫通領域が電位センサードメインであることは発見されていなかった.電位センサードメインを持つことに気がついた時点ですぐに電位依存性ホスホイノシチドホスファターゼという特性が連想されてもよかったのであるが,当時,電位センサードメインはイオンチャネルに固有の構造と考えられていた.ただ,電位センサードメインの動きについては,電位依存性チャネルの長い研究の歴史の中で,膜電位変化に伴って膜内でアミノ酸の電荷が移動することによる一過的な電流,いわゆるゲート電流を計測する手法が確立していた.当時大学院生だった佐々木真理(現大阪医科薬科大学)が,この奇妙な分子をアフリカツメガエル卵母細胞に発現させ,電位依存性チャネルのゲート電流計測と同様な計測を試みたところ,見事なゲート電流が計測された(図1).酵素領域を欠失させてもこのゲート電流は観察され,また電位センサードメインの膜電位感知に最も重要な構造であるS4セグメント(3節参照)の電荷をキャンセルすると観察されなくなった.これらは,VSPがイオンチャネルと同様に膜電位感知機構を持つことを示すと同時に,これまでイオンチャネルにおいてポア領域と一体としてでしか機能しないと考えられてきた電位センサードメインが,それ単独で機能できることを初めて示した結果となった.これを契機にVSPの分子特性の解析をさらに進め,岩崎広英(現 群馬大学)がPTEN様領域の酵素特性をin vitro malachite green assayにより明らかにし,村田喜理(現 大阪大学)がPI(4,5)P2依存的イオンチャネル活性や,PIPsと選択的に結合するpleckstrin homology domainを持つ蛍光タンパク質を用いて,VSPが電位依存的なPI(4,5)P2脱リン酸化活性を持つことを証明した1, 4).
ところで,ホヤ由来のVSPは発現系として用いたアフリカツメガエル卵母細胞との相性がよく大きなゲート電流が計測されたが,ほぼ同時期に海外の研究グループが哺乳類由来のVSP(当時TPTEまたはTPIPと呼ばれて機能未知の分子として論文も出されていた5, 6))の解析をしていたようである.哺乳類由来VSPは発現系細胞での細胞膜への発現量が不十分でゲート電流が計測できなかった(いまでも発現実験が困難であり,ホヤ由来VSPはどのオルソログ分子よりもアフリカツメガエル卵母細胞での発現量が多い).もしホヤ由来のVSPに着目していなかったら論文を発表できなかったかもしれない.またカタユウレイボヤのゲノム情報からは,他に電位依存性プロトンチャネルHv1[別名VSOP(voltage sensor domain only protein)]7, 8)や,ニューロン特異的に発現する電位センサードメイン分子TMEM266も同定された9).特に前者は,哺乳類ゲノムでコードされる分子量最小のイオンチャネルで,血球細胞での活性酸素産生の調節因子であり,またヒト精子の機能に関わる分子であることから,国内外で注目され多くの研究者により研究されている.
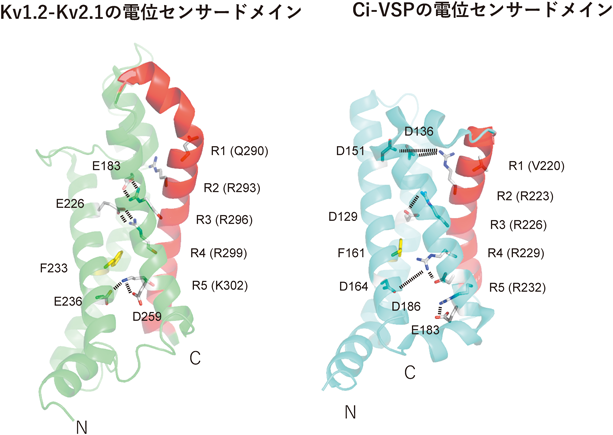
VSP遺伝子は棘皮動物からヒトまでほとんどの後口動物に保存され,前口動物では一部の動物種に限られている(たとえば線虫やショウジョウバエにはない)10).これまでにカタユウレイボヤ,ゼブラフィッシュ11),カスミサンショウウオ12),アフリカツメガエル13),ニワトリ14)のVSPの分子特性が明らかにされ,いずれも,脱分極でPI(4,5)P2を脱リン酸化する性質が明らかになっている.タンパク質構造は,ホヤ由来のVSPについて,電位センサードメイン15)と,細胞質領域(PTEN様領域)16, 17)が,別々のポリペプチドとして,X線結晶構造解析により明らかにされている.電位センサードメインはイオンチャネルにおけるそれと同様に四つの膜貫通領域からなり,4番目のS4セグメントに複数の塩基性アミノ酸が三つ置きに並び,他のヘリックスの酸性アミノ酸と塩橋を形成する(図3)15).細胞質領域はX線結晶構造で明らかにされているヒトPTEN18)と同様に,主にαヘリックスからなるホスファターゼドメインと膜と相互作用するC2ドメインから構成される16).ホスファターゼドメインには,多くのPTPase(プロテインホスファターゼ)に保存されるHCxxGxxR[またはC(x)5R]コンセンサス配列を持ち,特に活性中心のシステインは脱リン酸化反応に必須である19).電位センサードメインとホスファターゼドメインの間は塩基性アミノ酸を多く含む,約20アミノ酸からなるリンカー(VSD-PDリンカー)で結合されている.
1)電位センサードメインの性質
上述のように,アフリカツメガエル卵母細胞などの発現系を用いて,イオンチャネルの膜電位を感知する機能は,電位センサードメインの構造変化に伴う電流(ゲート電流)として,定量的に解析されてきた1).各膜電位に脱分極してゲート電流を計測し,移動した電荷総量を各電位についてプロットすることで,電位依存性の定量的な比較が容易になる.このような解析を行うと,VSPでは,電位依存性チャネルの場合よりも高い脱分極にしないと電位センサーが動かない.また1分子あたりに移動する電荷数は1から2程度であり1),一つの電位センサードメインあたり,3から4程度の電荷数が動く典型的な電位依存性チャネルに比較すると,「緩やかな」電位依存性を示す.
2)酵素活性の性質
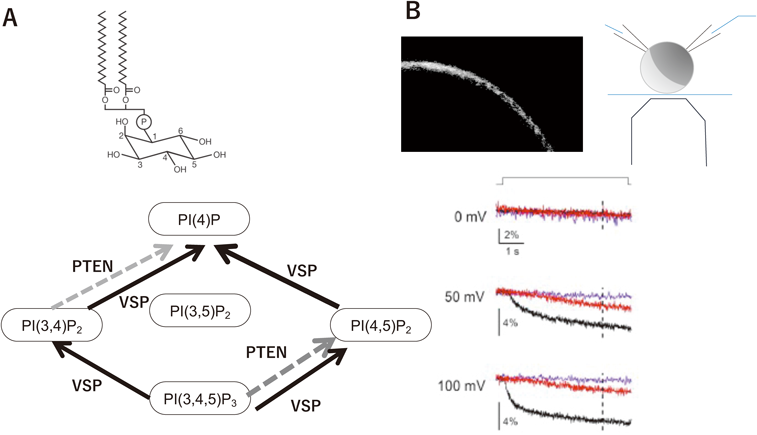
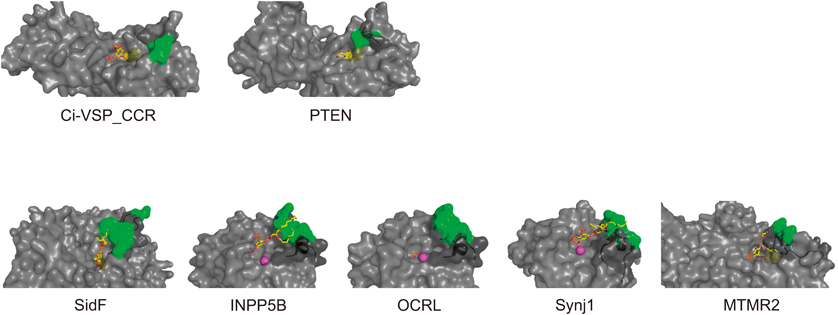
細胞質領域は脱リン酸化活性を示し,PTEN20)と同様,基質はタンパク質ではなく,膜脂質分子のホスホイノシチドである.大腸菌に発現させて精製した細胞内領域のみからなるポリペプチドについて,in vitro malachite green assayによりリン酸の遊離を計測すると,脱リン酸化反応のターンオーバー速度はPIP3に対して約2.2秒,PI(4,5)P2に対して,約3.6秒である16)(ただし,これは膜貫通領域を含まない部分配列での酵素活性である.細胞膜に組み込まれたVSPでの酵素活性は,in vitro malachite green assayでは計測されていない.細胞からのPI(4,5)P2特異的FRET蛍光計測結果から数理モデルを介して脱分極状況でのターンオーバー速度の最大値が推定されており21),PI(4,5)P2に対して約0.09秒であり,細胞内領域のポリペプチドについてin vitro malachite green assayで計測したターンオーバー速度の約40倍である).VSPは3位のリン酸に特異的活性を示すPTENとは異なり,PI(4,5)P2に対して5位のリン酸を脱リン酸化する基質特異性を持つ.また3位のリン酸の脱リン酸化活性も持ち合わせており,3種類のホスホイノシチド分子種に対して四つの反応を示す(図4A).このPTENとは異なる基質特異性の構造基盤について現在までわかっているのは,基質結合部位のアミノ酸(VSPの364Glyに対応する部位がPTENではAla)22)と,VSPには存在するがPTENにはみられないとされる,基質結合部位に覆いかぶさるループ状の構造(411Gluを含む数アミノ酸)17)である.これら二つの部位は,VSPオルソログ間で完全に保存されている.PTENにおいてこれらの部分をVSPのアミノ酸に変化させると,VSPのようにPI(4,5)P2の5位を脱リン酸化する活性を示すようになる23).一方で,逆の転換,つまりVSPにPTEN様の基質特異性を持たせることには成功していない.多くのタンパク質で膜への結合に関わるとされているC2ドメインは,VSPではあまり役割が明らかではない.PTENではC2ドメインの下流に,リン酸化を受ける領域があり,PTEN活性の調節やタンパク質分解に重要であることが知られている24).しかしVSPではこのC2ドメイン下流の領域は存在しない.
3)サブユニット構成
当初アフリカツメガエル卵母細胞発現系での一分子計測から,VSPは単量体と考えられた25)が,後に同じ研究者により,高い発現密度の状態では二量体となる26)ことが報告された.また酵素活性について単量体での協働性がある可能性が提示されたが反論する論文が出される27)など,現在まで二量体化の意義は明確ではない.
VSPにおける最大の謎は,いうまでもなくどうやって電位センサードメインの動きから酵素の活性化につながるのか,という点である.電位依存性チャネルでは,電位センサードメインは,膜内の膜貫通領域からなるポアドメインの構造変化を調節する.つまり膜貫通領域が他の膜貫通領域を調節する.電位センサードメインとポアドメインとの共役はイオンチャネル研究の重要なテーマであり,ポアドメインヘつながるリンカー部分(S4–S5リンカー)が,電位センサードメインのS4セグメントの動きに伴って機械的に動くことでポアドメインのヘリックスに構造変化が生じる,というモデルを中心に,現在も熱い論争が続いている.一方VSPにおけるエフェクターは細胞内領域の構造である.どのように電位センサードメインの構造変化が酵素を活性化するのかは,VSPに固有の問題としてだけでなく電位依存性膜タンパク質の動作原理全般を理解する上でも重要である.
1)電気化学共役
これまで電位感知機能と酵素活性との関係が,アフリカツメガエル卵母細胞などで膜電位を印加した状況で,ホスホイノシチドのレベルを,PI(4,5)P2感受性のイオンチャネルや,ホスホイノシチド選択的に結合する蛍光センサータンパク質を用いて定量する(図4)ことで明らかにされてきた19, 28, 29).これにより,VSPの電位センサードメインが示す幅広い膜電位範囲の一部だけで酵素活性が変化するというわけではなく,膜電位範囲全般にわたって酵素活性が連続的に変化する特性があることが明らかにされた.このことは,電位センサードメインと酵素領域が強く共役していることも示している.またこの共役は一方向ではなく,双方向である.実際,酵素領域のさまざまな部位にアミノ酸変異を導入すると,程度の違いはあるが,多くの変異において,電位センサードメインの動きのキネティックスが変化する10).
しかし詳細な仕組みの理解には,最終的な酵素活性の結果としてのホスホイノシチドのレベルをモニターするだけでなく,電位センサードメインの動きに伴う酵素領域のタンパク質構造の変化を捉えることが必要である.この目標に強力なツールとなったのが,非天然蛍光アミノ酸を用いた手法である.
蛍光性非天然アミノ酸の一種Anap[3-(6-acetylnaphthalen-2-ylamino)-2-aminopropanoic acid]は,構造がprodan[6-propionyl-2-(N,N-dimethyl)aminonaphthalene]と類似し,疎水性,親水性環境の変化で波長特性や蛍光強度が変化する.mRNAからの翻訳時にamber suppressor codonを認識して組み込むための直交のtRNAとtRNA合成酵素のペアが創製されている30)ので,これを用いて任意のアミノ酸部位に組み込むことが可能である.大きさもトリプトファン程度なので,GFPなどの大きな分子を組み込んだ場合とは違って局所の構造に干渉する懸念も少ない.坂田宗平(現 大阪医科薬科大学)は,AnapをVSPの細胞質領域のさまざまな部位に導入し,電位依存的に蛍光シグナルの変化を計測することで,局所の構造変化を捉えることに成功した31).その結果,ほとんどの部位が電位センサーの動きに依存し,ほぼ時間遅れなく局所の構造変化が生じることが明らかになった.このことは,酵素領域の一部だけが構造変化して酵素活性が出るのではなく,酵素領域全般に構造変化が生じることを示す.ちなみに,そのころMinorらのグループはカタユウレイボヤ由来VSP(Ci-VSP)の細胞質領域のX線結晶構造解析から基質結合部位付近の局所構造が異なる2種類の構造を得て,基質がアクセスする結合ポケットのサイズが変化することで,酵素活性が調節される,とする17)興味深い説を提示したが,我々のAnapによる結果はこのモデルを支持しない.
さらに,Anapの蛍光波長と重なる吸収スペクトルを持つDipicrylamine(DPA)をFRETの対として利用することで膜と酵素領域の間の距離の変化を調べたところ,C2ドメインと膜との距離は変化しないことが明らかになった(ただしこの計測は現在のところC2ドメインの1か所の部位でのみ行われており,電位センサードメインに近い酵素ドメインでは細胞膜との距離の計測はできていない).
2)ホスホイノシチドホスファターゼ共通の制御スイッチがある?
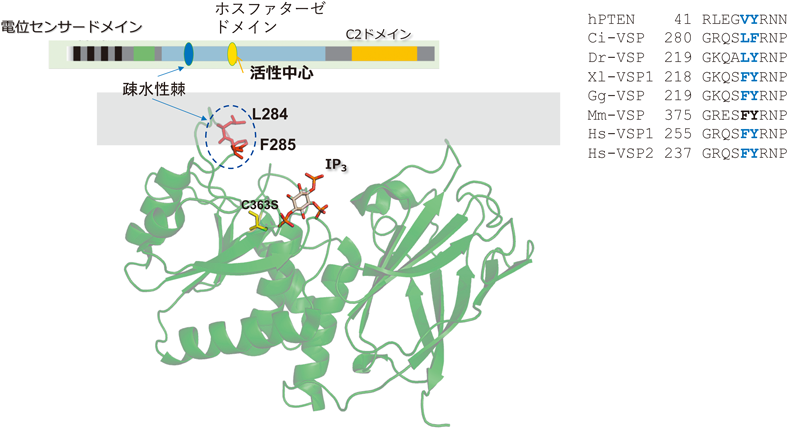
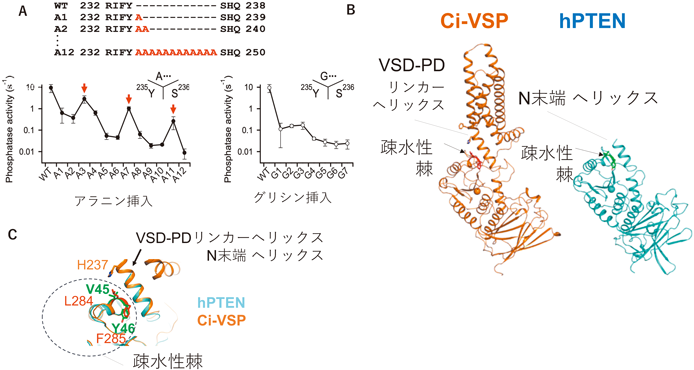
西澤和久ら(当時帝京大学)はVSPの細胞質領域のX線結晶構造を元に,MDシミュレーションで酵素領域が膜にどのように相互作用するかを解析した結果,VSPの酵素ドメインに,膜に突き刺さる疎水性部分(Ci-VSPの番号で284Phe, 285Leu)の存在に気がついた[西澤博士はこれを疎水性棘(hydrophobic spine)と呼んだ](図5).この部分はほとんどの種のVSPオルソログに保存されていた.興味深いことにさまざまなホスホイノシチドホスファターゼの立体構造を調べると,PTENをはじめ多くの分子で基質結合ポケットの近傍に,VSPの疎水性棘と同様な構造がみられた(図6).この部位の役割を明らかにするため,川鍋陽(現 香川大学)は,284Pheと285Leuに変異を導入し,酵素活性を計測した.これらは酵素活性が顕著に減弱しており,興味深いことにPTENにおける相同な部位の変異においても活性が減弱していた.さらにAnapを用いて酵素領域の動きを解析したところ,野生型VSPではAnap蛍光変化の速い変化と遅い変化の二つの成分があるが,変異体では,遅い成分が消失することを見いだした.このことから,疎水性棘には,電位センサーから酵素活性の伝達の過程で酵素活性を発揮する際のスイッチのような役割があると考えられた32).
3)疎水性棘は,電気化学信号変換のハブである
次の疑問は,疎水性棘はどうやって電位センサードメインの動きで制御されるかという点である.電位センサードメインのS4セグメントの構造をオルソログ分子間で比較すると,VSPのS4セグメントには電位を感知する塩基性アミノ酸が規則的に並んでいるが,最も下部の塩基性アミノ酸(231Arg)の直下に二つの疎水性アミノ酸(Ci-VSPでは234Phe, 235Tyr)が保存されていることがわかった.この部位を変異させると,電位センサーの動きはあまり影響がないにもかかわらず,酵素活性が大きく減弱し,この部位がカップリングに重要であることが示された.AlphaFold2を用いて全長タンパク質の立体構造予測を行った結果,S4セグメント直下から細胞質側へ続く領域[以下,voltage sensor domain-phosphatase domain(VSD-PD)リンカー]は上記の234Pheと235Tyrを含めて,S4セグメントから連続する長いヘリックス構造をとっていた.一方,234Pheの直後にさまざまな長さのアラニンを挿入した分子を作製して電位依存的酵素活性を計測すると,挿入するアラニンの長さに依存して周期的に酵素活性が変化し,その周期がαヘリックスの周期に近い数字(3または4アミノ酸)であったことからも,VSD-PDリンカーはヘリックス構造をとると考えられた(図7A).
S4セグメント直下の構造が疎水性であることから,ここが酵素ドメインの疎水性棘と直接相互作用するのではないかと想定し,トリプトファンによるAnap蛍光のクエンチング 法,および,システイン導入によるクロスリンキングの検証を行った.その結果VSD-PDリンカーは膜電位依存的に疎水性棘と近接することが明らかになった.AlphaFold2での構造予測でも,この疎水性棘がVSD-PDリンカーのヘリックスとほぼ直角に近接する構造がみられた33).これらの知見から,膜電位の脱分極により,S4セグメント直下のヘリックスと酵素ドメインの疎水性棘の,二つの疎水性のヘリックスどうしの相互作用を介して「電位センサードメインのS4セグメントの上方向の動き→疎水性棘の構造変化(膜への移動)→酵素の活性化」が生じると考えられる.この疎水性構造どうしの相互作用は,電位依存性K+チャネルのKv2.1のゲーティング機構として最近発見されたhydrophobic nexusと呼ばれる,S4セグメント下流の疎水性構造とエフェクターであるポアドメインのヘリックスの疎水性アミノ酸との相互作用34)を連想させるものである.さらに,VSPのS4セグメントが細胞膜を貫通して細胞質側へ長くヘリックスを伸ばしている構造は,最近低温電子顕微鏡解析で明らかにされた,精子特異的ナトリウムプロトン交換体[sperm specific sodium proton exchanger(sNHE)またはSLC9C1]の構造35)においてS4セグメントから細胞質側の環状ヌクレオチド結合ドメインに向けて連続して長く伸びたヘリックス構造と類似している.電位センサードメインが,細胞質側の構造変化をもたらすこれら2種の分子間で類似した特徴を示している点は,とても興味深い.
4)PTENの酵素活性調節との関連
PTENにおいてN末端側はαヘリックス構造をとっている.この部位はPTENの酵素活性に必須であり,この領域での点変異が発がんや悪性化につながることがよく知られている.PTENの全長の構造をAlphaFold2で予測すると,N末端ヘリックスはVSPにおけるVSD-PDリンカーヘリックスと重なり,さらに疎水性棘もほぼ同じ位置に存在する(図7).このことから,VSPにおけるS4セグメントの動きによる酵素活性調節は,もともとPTENに内在されている,N末端側のヘリックスと疎水性棘とのhelix to helix相互作用による調節機構を利用した仕組みとして理解できるのかもしれない.
5)基質特異性が電位センサードメインの状態で変化するのか
VSPはPI(4,5)P2の5位のリン酸を脱リン酸化する活性が主であるものの,弱いながらPIP3やPI(3,4)P2の3位のリン酸の脱リン酸化活性も持ち合わせている36).PI(3,4)P2特異的な蛍光プローブを用いて膜電位固定下で膜電位を印加すると,PI(3,4)P2は最初増加したあとに減少するという二相性の変化を示す.最初の増加は,PIP3の5位の脱リン酸化によるもので,次にみられる減少はPI(3,4)P2の3位の脱リン酸化によるものである.この変化は印加する膜電位のレベルによって異なり,増加の変化は低い膜電位への脱分極で観察されるのに対して,PI(3,4)P2レベルの減少は高い脱分極で顕著にみられた.Isacoffのグループは,複数の電位センサードメインの変異体を用いて,こうした酵素活性の電位依存性を解析し,低い電位ではPIP3が優先的に基質となり,高い電位ではPI(4,5)P2,PI(3,4)P2が優先的に基質となるというモデルを提唱した37).しかし計測が行われたアフリカツメガエル卵母細胞では電位印加前のホスホイノシチド濃度をコントロールすることは難しく,四つの反応(図4A)が複雑に重なっていて見かけ上基質特異性が膜電位に依存するようにみえている可能性がある.実際,Hilleらのグループは,HEK293T細胞で,各ホスホイノシチドレベルを蛍光プローブで系統的に計測した解析と数値シミュレーションを対照させる解析を行い,四つの反応の間で,反応速度の電位依存性は共通であると結論した.つまり,Isacoffらの考えとは異なり,電位センサードメインの状態により基質の選択にバイアスがかかることはないという結論である21).ただし,我々は疎水性棘の解析において酵素の活性化には単一の状態遷移でなく複数の遷移が関わることを明らかにしており28, 32),単一の遷移のみを想定しているHilleらのシミュレーションは単純化したものである.電位センサードメインの状態によってVSPの基質選択性が変化するかどうかはまだ決着がついていない.
では,こうしたVSPの分子特性は生体ではどのように活用されているのだろうか? VSPの動作原理の理解とは対照的に,その生物学的役割については長らく不明であった.最近モデル生物での研究が行われ,ようやく分子の役割が明らかになってきた.
1)精子でのVSPの機能
Ci-VSPが同定された際,EST(expressed sequence tag)の配列情報で精巣に発現が認められ,Ci-VSPに対する特異抗体を用いた免疫電子顕微鏡の解析により精子の鞭毛の細胞膜に発現することが明らかになった1).その後哺乳類については,河合喬文(現 大阪大学)が伊川正人らとVSPノックアウトマウス(新潟大学の崎村建司らが作製)を用いて精子の解析を行い,精子の形態には異常がないがin vitroでの受精率が極端に低いこと,また精子の成熟後の運動性に異常があることが見いだされた.精子は精巣管で形成され,精巣上体を移動し,射精のあと,卵管内で最終的に卵と受精するが,このためにはcapacitation(受精能獲得)と呼ばれる成熟過程が必要である.具体的には精子頭部での消化酵素の開口放出(先体反応)と,鞭毛の大きな角度での大きな運動(超活性化)である.VSPをノックアウトしたマウスの精子では,超活性化に異常が生じており,個々の精子の運動の軌跡をトラッキング解析してみると,同じところを回転する異常な動きを示した.一方,ホスノイノシチド成分について質量分析による解析を行ったところ,ノックアウトマウスの精子では野生型に比べPI(4,5)P2の量が増え,逆にPI(4)Pの量が減っていることが明らかになり[佐々木雄彦(現 東京医科歯科大学)との共同研究],VSPが精子でPI(4,5)P2を基質とする酵素として働いていることが確認された.
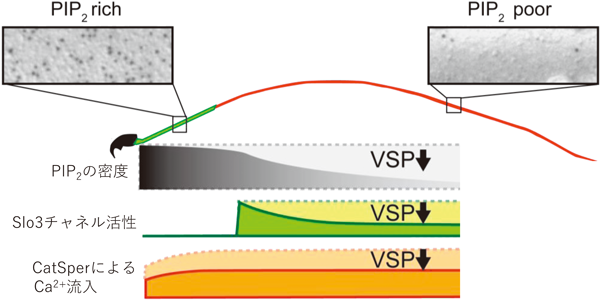
capacitation後の精子の運動性には,細胞外からのCa2+流入が重要な役割を果たすことが知られている.Ca2+イメージングの実験で,ノックアウトマウス由来の精子ではCa2+濃度の上昇が正常マウスのそれより顕著であり,またCaポンプの阻害剤では影響を受けないことから,Ca2+流入が亢進していると考えられた.パッチクランプ法によるイオン電流の計測で,精子のCa2+チャネルであるCatSperの活性は野生型と変わらないが,膜電位を過分極させる精子特異的カリウムチャネル電流(KSper)の量が増えていた.このKSperは受精に必須で38),分子実体としては,アルカリで活性化される電位依存性カリウムチャネル(Kvチャネル)であるSlo3チャネルにより担われている.Slo3チャネルはその活性にPI(4,5)P2を必要とすることが知られている.そこでVSPの酵素活性がPI(4,5)P2の変化を介してSlo3チャネルの活性に影響し,CatSperからのCa2+流入を増加させると考えるに至った8).
しかし,この段階で新たな疑問が生じた.ニューロンの神経伝達物質による興奮調節としてよく知られるKvチャネルの一種Mチャネル(KCNQ2/3複合体またはKv7.2/Kv7.3複合体)や,多様な物理的または化学的センシングに関わるTRPチャネルなどと異なり,Slo3チャネルはPI(4,5)P2結合親和性がきわめて高く,生理的にPI(4,5)P2の量の変動がSlo3チャネルの活性を実際に調節しているとは考えにくい.我々は,精子の鞭毛の膜でのPI(4,5)P2の空間的分布に鍵があるのではと考え,藤本豊士(当時名古屋大学)らの協力を仰ぎ,凍結レプリカによる脂質の局在を電子顕微鏡で解析する技術により,PI(4,5)P2の生体膜中での分布を検証した.その結果,精子の鞭毛の膜では,PI(4,5)P2は均一に分布しているわけでなく,鞭毛の先端ほど密度が低くなっていた.VSPをノックアウトした精子では,こうした鞭毛の前後軸でのPI(4,5)P2の密度の勾配はみられなかった.これらから,正常のマウスの精子では,細胞全体のPI(4,5)P2の平均密度でなく,鞭毛膜のPI(4,5)P2の空間分布様式が,実際にactiveなSlo3チャネルの数を規定しており,VSPはPI(4,5)P2の分布を精子尾部で調整することで,Slo3チャネル活性を調節して適度なCa2+流入を維持していると結論した8)(図8).
これらの結果は,精子の生物学としての新しい知見であるだけでなく,イオンチャネルの活性制御の点でも新しい発見である.これまで解析されてきたニューロンや筋のイオンチャネルでは,イオンチャネルの分子数よりPI(4,5)P2の分子数の方が多く,またPI(4,5)P2に対して適度に緩い結合親和性をもつことで,GPCRのシグナル伝達などによって生じるPI(4,5)P2の濃度変化に応じて細胞でのイオンチャネル活性が変化すると考えられてきた.一方,今回のVSPのKOマウスの解析から明らかになった仕組みでは,Slo3チャネルが高いPI(4,5)P2親和性をもつことと,疎にPI(4,5)P2が分布する(おそらくSlo3チャネル分子の数よりも少ないだろうと考えている)ことで,ホスホイノシチド分子の空間分布様式によってイオンチャネル活性が規定される.こうしたメカニズムが精子に固有なイオンチャネル制御の仕組みであるのかは,今後さらなる解析が必要であろう.
2)消化管上皮細胞での役割
分子の発見後,VSPが精子だけでなく,ニワトリの中腎の上皮細胞39)やカタユウレイボヤの幼若体の消化管上皮40)など,上皮細胞に発現することが明らかにされた.最近,ゼブラフィッシュにおいて抗体を用いた免疫染色とGFPノックイン系統の解析により,VSPは消化管後方の,受容体依存的エンドサイトーシス機能を持つ特殊な上皮細胞[Duke大学のBagnatらのグループによりlysosome-rich enterocyte(LRE)と命名されている41)]に顕著に発現することが示された42).哺乳類では消化管からの栄養吸収は,胃や十二指腸での細胞外での消化酵素による消化と,細胞膜での各種トランスポーター分子を介する選択的輸送により行われるが,魚類や無脊椎動物を含む多くの生物は,受容体依存的な食作用や飲作用により吸収して細胞内のリソソームで消化する原始的なシステムを併せ持っている.LREはこの後者のシステムを担う細胞である(ゼブラフィッシュなどの胃を持たない硬骨魚類では後者のシステムが特に発達している43)).ゼブラフィッシュの稚魚の消化管に強制的に蛍光デキストランやmCherryなどの蛍光タンパク質を投与して,共焦点蛍光顕微鏡によるライブイメージング観察を行ったところ,VSP遺伝子をノックアウトした稚魚ではこれら分子の吸収過程に異常がみられた.またVSPノックアウト個体では孵化後の生存率が有意に低下していた.またカタユウレイボヤ幼若体の消化管でも受容体依存的エンドサイトーシス過程に関わる遺伝子群とVSPが同じ上皮細胞に発現しており,上皮細胞でのVSPの機能は進化的にも保存されていると考えられる.
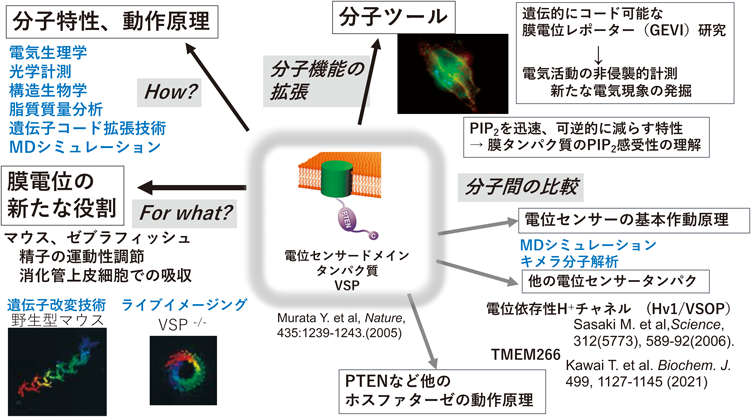
当初予想していなかった研究展開として,分子ツールとしての研究があげられる.
1)GEVI(genetically encoded voltage indicator)
VSPは,現在ロドプシン関連分子とともに,in vivoでの細胞活動を可視化するための,遺伝コード可能な膜電位プローブGEVIの開発の材料として用いられている.VSPの発見前には,電位依存性チャネルにGFPなどの蛍光タンパク質を融合させることでタンパク質性の膜電位プローブとなる可能性が示された44)が,発現量やシグナルの弱さなどの点で実用的でなかった.下流の酵素領域と切り離して単独で機能できるというモジュール性から,多くの研究者による多様な試行錯誤がなされるようになり,速い応答性と選択的な膜ターゲット効率を備えた実用性の高いプローブが開発されてきた45).現在2光子顕微鏡で実用的に膜電位変化のin vivoでの計測に使えるプローブのほとんどがVSPを活用したものになっている.
2)PI(4,5)P2を急激に,かつ可逆的に減らすツール
VSPは電位依存的にPI(4,5)P2ホスファターゼ活性を示すため,イオンチャネルのcDNAとともに培養細胞などに発現させれば,通常の電気生理学計測系をそのまま用いて実験が可能であり,イオンチャネル研究者にとっては特段の設備の追加の必要がない.そのためこれまで多くの研究者によりイオンチャネルや起電性トランスポーター分子の研究に活用されてきた45–47).ホスホイノシチドを短時間で一過的に減少させる能力は重要で,これが遅い方法,たとえばwortmanninなどを用いた薬理学的手法の場合には,直接の効果と,形態変化や膜トラフィックなどを介する二次的変化を区別しにくい.また,rapamycinを用いた分子会合を利用した酵素活性調節法であると,いったん分子会合により活性が誘導された酵素はそのまま活性を維持するので摂動を解除できず,一過的に制御可能なVSPとは対照的である.我々は,VSPの動作原理の理解に基づいて,ツールとしての性質を向上させる試みも行ってきた47)(Addgeneに登録).なお,基質特異性をPIP3特異的に変更したVSPについては,他のグループによりCi-VSPの電位センサーとPTENの酵素領域をキメラにした分子(VSPTEN48))が作られているが,電位依存性と迅速性の点で使いにくい面がある.
これまでのVSPの研究のインパクトは以下にまとめられる(図9参照).
- (1)電位センサードメインが,従来知られていたイオンチャネルのイオン透過機構以外の生理機能(ホスホイノシチドの脱リン酸化反応)に使われることの発見.
- (2)電位センサードメインが独立した機能モジュールであることの発見とその応用(GEVIの開発での利用など).
- (3)電位センサードメインの動作原理研究のためのシンプルなモデルを提示.
- (4)がん化やがんの悪性化に重要なPTENの構造機能連関の理解への端緒を提供.
- (5)精子の運動性調節におけるホスホイノシチドによるイオンチャネル(Slo3)の調節の発見,また消化管上皮でのエンドサイトーシス依存的栄養吸収に関わる新規のシグナルの可能性.
- (6)細胞膜のPI(4,5)P2を減少させるツールとしての有用性[迅速かつ可逆的にPI(4,5)P2を減らす特性ゆえ,多くのイオンチャネルの脂質分子による調節の研究に用いられるようになった].
以下に今後の課題と展望を述べる.
1)VSPの電気化学連関の機構
上述のように膜電位変化により酵素領域と電位センサードメイン間のリンカーヘリックスが,酵素領域の疎水性棘と相互作用することが,電気化学連関機構の中心であると考えられる.しかし,静止状態の構造からの遷移が,膜電位変化によりどのように起こるのかは未解明である.疎水性棘がどのように酵素活性を調節するのかも詳細は明らかでない.今後の進展には構造生物学と計算化学によるアプローチが不可欠であろう.最近MacKinnonらにより,リポソーム膜に膜電位を印加した状況でKvチャネルの構造が解かれた49).またこれまで分子量が小さくて低温電子顕微鏡による単粒子解析の対象にはなりにくかった分子でも構造が解けるようになりつつあり,今後VSPについて活性化状態と静止状態の構造情報が得られることで動作原理の解明が進むと期待される.VSPが活性化した際にS4セグメントの下流(VSD-PDリンカー)が長いヘリックス構造を示す一方,電位センサードメインが細胞内側へ沈み込んでいる静止状態(膜電位が過分極した状態)では,この部位がどのような構造をとっているか,という疑問がある.最近,我々はAnap蛍光のvoltage clamp fluorometry解析により,過分極で生じる電位センサードメインの動きは,脱分極により生じる動きの場合に酵素領域からの逆向性の影響がみられるのとは対照的に,こうした酵素領域からの影響がみられないことを発見した50).このことはS4セグメントの下流のヘリックス構造は過分極時には異なる構造,たとえば,ヘリックスがほどけた状態にある可能性を示すのかもしれない.なお,電位センサードメインが静止状態での構造との関連で,もう一つ重要な未解決の問題がある.それは,静止状態において酵素活性が完全に消失しているのか,それとも弱いながらも潜在的な活性があるのかという点である.
またホスホイノシチドの酵素全般に共通する問題として,活性中心にどのようにホスホイノシチドが結合し,脱リン酸化後にあらたな基質と入れ替わるか,という点も今後の課題であろう.活性中心のシステイン(Ci-VSPでは363Cys)の位置にホスホイノシチドのリン酸が結合するには,膜からホスホイノシチドが細胞内側に下がる必要があると考えられる.基質が活性中心にドッキングする経路と,反応物[PI(4,5)P2が基質の場合はPI(4)P]が外れて膜に戻っていく経路や環境が同じなのだろうか.これらの問いに答えるには,今後構造情報とともに,膜の動態を含めた計算化学的なアプローチが重要と思われる.
2)PTENとの共通性
VSPの電位依存的活性が見いだされる前には,VSPはPTEN関連酵素として記載され,単にPTENが膜にアンカーされた酵素のような分子と考えられていた.前述したように,疎水性棘が両者に共通であったり,PTENのN末端のヘリックス構造がVSPのVSD-PDリンカー部と類似していたりする点などからも,VSPの酵素調節機構には,PTENのそれと共通の仕組みがあると考えられる.しかし,VSP=電位センサードメイン(膜貫通領域)+PTENという単純な構図は正しくない.まずVSPの基質特異性をPTENの性質に変更することが実験的に成功しておらず,活性中心のアミノ酸変異(G364A変異)により電位依存的酵素活性が大きく減弱することが知られている.また最近,有袋類のVSPの多くが基質結合部位のコンセンサス配列のC(x)5RのCysの一つC末端側のアミノ酸が多くのVSPでみられるLysではない点に着目して,Ci-VSPにおいて,この部位を多様なアミノ酸に置換したところ,基質特異性には大きな変化が生じず,むしろ電位センサードメインとのカップリングに影響が出た51).このことは,進化の過程で,電位センサードメインと酵素領域が融合しただけでなく,酵素が電位センサードメインでうまく調節されるためには酵素領域内も改変される必要があったことを物語っている.そのため,VSPにおいて電位依存性を持つことと,PTENと異なる基質特異性を有することは完全に切り離して考えることができない可能性が大きい.PTENでは,PI(4,5)P2がN末端側構造に結合することで酵素活性が調節されると考えられてきた52).結合部位のアミノ酸配列はVSPでも保存されているが,実際にPI(4,5)P2による酵素活性による調節がVSPにも存在するのかは,かつてこれを示唆する論文が出されたものの53, 54),現在まで決着がついていない.難点の一つはPI(4,5)P2はVSPの基質であり,調節因子としてのPI(4,5)P2と基質としてのPI(4,5)P2を区別して解析することが困難である点である.
3)VSPの生理機能
精子や消化管上皮で,VSPが,シグナル伝達ネットワークのどこにはまるか?という問題に答えるためには,in vivoでVSPがどのような刺激により活性化されているのかを知ることが重要である.これにはinputとなる刺激(たとえば膜電位変化)を精子や上皮などで計測することが課題となる.イオンチャネルをはじめとする膜機能分子の構造活性相関は,これまで構造解析と発現系細胞での機能計測などにより理解が飛躍的に進んだ一方,分子特性と生理現象との間には依然大きなギャップが残されてきた.これまでの電気生理学的技術は,多くの革新がなされてきたものの電極をアプローチする際の物理的制約があって計測できる対象が限定されてきた.膜電位とかpHなど,物理や化学では古くから研究されてきた基本的なパラメーターについて,in vivoでの動態はいまだに謎に包まれている.体内奥深く存在する臓器から細胞レベルでin vivoでの状況を調べるには,新たな技術の創製が必要であろう.
4)機能モジュールとしての電位センサードメイン(他の電位センサードメインタンパク質との共通性,多様性)
VSPの発見は,電位センサードメインが,かつて考えられたような電位依存性チャネルに固有の分子内構造でなく,独立した機能モジュールであることを明示し,さらに電位センサードメインのみからなる分子でチャネル機能を持つ分子(Hv1またはVSOP)や,ニューロンに発現する機能未知の膜タンパク質(TMEM266)の発見につながった.電位センサードメインが二次性トランスポーター機能を電位依存的に調節するsNHE(SLC9C1)なども含めて,これまで想像されていた以上に電位センサードメインには柔軟なポテンシャルがあり多様な生理機能に使われていることが明らかになってきた.これら分子間を相互に比較することで,電位センサードメインの基本動作原理が解明されるとともに,分子の改変によって生命がどのように,したたかに環境に適応してきたのかが,はっきりとみえてくると期待したい.これらの分子が創生されてきた時間関係も興味深い.進化の過程で単独の電位センサードメインがまず先に創生され,他のモジュール構造と組み合わさったのだろうか? もしそうだとすると,地球史上最初の電位センサードメインの機能はHv1/VSOPのようなチャネル機能であったのだろうか? それとも違う機能があったのだろうか? 祖先の生物でのゲノム中でエキソンのシャッフリングがどのように生じ,新たなハイブリッド的分子機能が創生されたのかも興味深い問いである.ちなみに,上記にあげた分子のうち,TMEM266以外はすべてヒトでは精子特異的に発現している.精子のCa2+チャネル(CatSper)やK+チャネル(Slo3)も精子特異的な発現を示す分子であることも合わせて,精子が多様な膜電位依存性分子を発現している点は興味深い.
VSPの研究はこれまで述べてきたように多面的であるが,それは研究が進むにつれて枝葉が分かれるように広がった結果である.一方で分子,細胞,個体レベルと高階層を横断して理解が進むことで,個体レベルでの機能の視点から,分子特性の理解が深まり,逆に,分子特性を深く知ることで個体レベルでの新たな仕組みがみえるのではないだろうか.たとえば,もし電位センサードメインの状態によって基質特異性が変化するとされる分子特性(前述)が事実であれば,実際に細胞の膜電位の到達レベルや数秒間の履歴により細胞内のホスホイノシチドの存在比や分布様式が変化し,それが生理的機能に結びついている可能性がある.その一方,膜電位プローブなど,分子をいったん個体での機能から切り離してタンパク質のモジュール性に基づいて新たな特性を創造できる点も重要であろう.今後も多面的に研究が進み,さらに「新たな世界が視える」ことを期待したい.
謝辞Acknowledgments
図の作製で協力いただいた水谷夏希博士に感謝いたします.本稿で述べた研究のうち著者が関わった内容は,すべて多くの共同研究者の研究活動とサポートスタッフによる厚い協力によりなされたものです.共同研究者,ラボのOB, OG, 現在のメンバーに深く感謝いたします.
引用文献References
1) Murata, Y., Iwasaki, H., Sasaki, M., Inaba, K., & Okamura, Y. (2005) Phosphoinositide phosphatase activity coupled to an intrinsic voltage sensor. Nature, 435, 1239–1243.
2) Dehal, P., Satou, Y., Campbell, R.K., Chapman, J., Degnan, B., De Tomaso, A., Davidson, B., Di Gregorio, A., Gelpke, M., Goodstein, D.M., et al. (2002) The draft genome of Ciona intestinalis: Insights into chordate and vertebrate origins. Science, 298, 2157–2167.
3) Okamura, Y., Nishino, A., Murata, Y., Nakajo, K., Iwasaki, H., Ohtsuka, Y., Tanaka-Kunishima, M., Takahashi, N., Hara, Y., Yoshida, T., et al. (2005) Comprehensive analysis of the ascidian genome reveals novel insights into the molecular evolution of ion channel genes. Physiol. Genomics, 22, 269–282.
4) Murata, Y. & Okamura, Y. (2007) Depolarization activates the phosphoinositide phosphatase Ci-VSP, as detected in Xenopus oocytes coexpressing sensors of PIP2. J. Physiol., 583, 875–889.
5) Walker, S.M., Downes, C.P., & Leslie, N.R. (2001) TPIP: A novel phosphoinositide 3-phosphatase. Biochem. J., 360, 277–283.
6) Chen, H., Rossier, C., Morris, M.A., Scott, H.S., Gos, A., Bairoch, A., & Antonarakis, S.E. (1999) A testis-specific gene, TPTE, encodes a putative transmembrane tyrosine phosphatase and maps to the pericentromeric region of human chromosomes 21 and 13, and to chromosomes 15, 22, and Y. Hum Genet., 105, 399–409.
7) Sasaki, M., Takagi, M., & Okamura, Y. (2006) A voltage sensor-domain protein is a voltage-gated proton channel. Science, 312, 589–592.
8) Kawai, T., Miyata, H., Nakanishi, H., Sakata, S., Morioka, S., Sasaki, J., Watanabe, M., Sakimura, K., Fujimoto, T., Sasaki, T., et al. (2019) Polarized PtdIns(4,5)P2 distribution mediated by a voltage-sensing phosphatase (VSP) regulates sperm motility. Proc. Natl. Acad. Sci. USA, 116, 26020–26028.
9) Kawai, T., Narita, H., Konno, K., Akter, S., Andriani, R.T., Iwasaki, H., Nishikawa, S., Yokoi, N., Fukata, Y., Fukata, M., et al. (2022) Insight into the function of a unique voltage-sensor protein (TMEM266) and its short form in mouse cerebellum. Biochem. J., 479, 1127–1145.
10) Okamura, Y., Kawanabe, A., & Kawai, T. (2018) Voltage-sensing phosphatases: Biophysics, physiology, and molecular engineering. Physiol. Rev., 98, 2097–2131.
11) Hossain, I., Iwasaki, H., Okochi, Y., Chahine, M., Higashijima, S., Nagayama, K., & Okamura, Y. (2008) Enzyme domain affects the movement of the voltage sensor in ascidian and zebrafish voltage-sensing phosphatases. J. Biol. Chem., 283, 18248–18259.
12) Mutua, J., Jinno, Y., Sakata, S., Okochi, Y., Ueno, S., Tsutsui, H., Kawai, T., Iwao, Y., & Okamura, Y. (2014) Functional diversity of voltage-sensing phosphatases in two urodele amphibians. Physiol. Rep., 2, e12061.
13) Ratzan, W.J., Evsikov, A.V., Okamura, Y., & Jaffe, L.A. (2011) Voltage sensitive phosphoinositide phosphatases of Xenopus: Their tissue distribution and voltage dependence. J. Cell. Physiol., 226, 2740–2746.
14) Yamaguchi, S., Kurokawa, T., Taira, I., Aoki, N., Sakata, S., Okamura, Y., & Homma, K.J. (2014) Potential role of voltage-sensing phosphatases in regulation of cell structure through the production of PI(3,4)P2. J. Cell. Physiol., 229, 422–433.
15) Li, Q., Wanderling, S., Paduch, M., Medovoy, D., Singharoy, A., McGreevy, R., Villalba-Galea, C.A., Hulse, R.E., Roux, B., Schulten, K., et al. (2014) Structural mechanism of voltage-dependent gating in an isolated voltage-sensing domain. Nat. Struct. Mol. Biol., 21, 244–252.
16) Matsuda, M., Takeshita, K., Kurokawa, T., Sakata, S., Suzuki, M., Yamashita, E., Okamura, Y., & Nakagawa, A. (2011) Crystal structure of the cytoplasmic phosphatase and tensin homolog (PTEN)-like region of Ciona intestinalis voltage-sensing phosphatase provides insight into substrate specificity and redox regulation of the phosphoinositide phosphatase activity. J. Biol. Chem., 286, 23368–23377.
17) Liu, L., Kohout, S.C., Xu, Q., Müller, S., Kimberlin, C.R., Isacoff, E.Y., & Minor, D.L. Jr. (2012) A glutamate switch controls voltage-sensitive phosphatase function. Nat. Struct. Mol. Biol., 19, 633–641.
18) Lee, J.O., Yang, H., Georgescu, M.M., Di Cristofano, A., Maehama, T., Shi, Y., Dixon, J.E., Pandolfi, P., & Pavletich, N.P. (1999) Crystal structure of the PTEN tumor suppressor: Implications for its phosphoinositide phosphatase activity and membrane association. Cell, 99, 323–334.
19) Okamura, Y. & Dixon, J.E. (2011) Voltage-sensing phosphatase: Its molecular relationship with PTEN. Physiology (Bethesda), 26, 6–13.
20) Maehama, T. & Dixon, J.E. (1998) The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate. J. Biol. Chem., 273, 13375–13378.
21) Keum, D., Kruse, M., Kim, D.I., Hille, B., & Suh, B.C. (2016) Phosphoinositide 5- and 3-phosphatase activities of a voltage-sensing phosphatase in living cells show identical voltage dependence. Proc. Natl. Acad. Sci. USA, 113, E3686–E3695.
22) Iwasaki, H., Murata, Y., Kim, Y., Hossain, M.I., Worby, C.A., Dixon, J.E., McCormack, T., Sasaki, T., & Okamura, Y. (2008) A voltage-sensing phosphatase, Ci-VSP, which shares sequence identity with PTEN, dephosphorylates phosphatidylinositol 4,5-bisphosphate. Proc. Natl. Acad. Sci. USA, 105, 7970–7975.
23) Leitner, M.G., Hobiger, K., Mavrantoni, A., Feuer, A., Oberwinkler, J., Oliver, D., & Halaszovich, C.R. (2018) A126 in the active site and TI167/168 in the TI loop are essential determinants of the substrate specificity of PTEN. Cell. Mol. Life Sci., 75, 4235–4250.
24) Dempsey, D.R., Viennet, T., Iwase, R., Park, E., Henriquez, S., Chen, Z., Jeliazkov, J.R., Palanski, B.A., Phan, K.L., Coote, P., et al. (2021) The structural basis of PTEN regulation by multi-site phosphorylation. Nat. Struct. Mol. Biol., 28, 858–868.
25) Kohout, S.C., Ulbrich, M.H., Bell, S.C., & Isacoff, E.Y. (2008) Subunit organization and functional transitions in Ci-VSP. Nat. Struct. Mol. Biol., 15, 106–108.
26) Rayaprolu, V., Royal, P., Stengel, K., Sandoz, G., & Kohout, S.C. (2018) Dimerization of the voltage-sensing phosphatase controls its voltage-sensing and catalytic activity. J. Gen. Physiol., 150, 683–696.
27) Kruse, M., Kohout, S.C., & Hille, B. (2019) Reinterpretation of the substrate specificity of the voltage-sensing phosphatase during dimerization. J. Gen. Physiol., 151, 258–263.
28) Sakata, S. & Okamura, Y. (2014) hosphatase activity of the voltage-sensing phosphatase, VSP, shows graded dependence on the extent of activation of the voltage sensor. J. Physiol., 592, 899–914.
29) Sakata, S., Hossain, M.I., & Okamura, Y. (2011) Coupling of the phosphatase activity of Ci-VSP to its voltage sensor activity over the entire range of voltage sensitivity. J. Physiol., 589, 2687–2705.
30) Wang, L., Xie, J., & Schultz, P.G. (2006) Expanding the genetic code. Annu. Rev. Biophys. Biomol. Struct., 35, 225–249.
31) Sakata, S., Jinno, Y., Kawanabe, A., & Okamura, Y. (2016) Voltage-dependent motion of the catalytic region of voltage-sensing phosphatase monitored by a fluorescent amino acid. Proc. Natl. Acad. Sci. USA, 113, 7521–7526.
32) Kawanabe, A., Hashimoto, M., Nishizawa, M., Nishizawa, K., Narita, H., Yonezawa, T., Jinno, Y., Sakata, S., Nakagawa, A., & Okamura, Y. (2018) The hydrophobic nature of a novel membrane interface regulates the enzyme activity of a voltage-sensing phosphatase. eLife, 7, e41653.
33) Mizutani, N., Kawanabe, A., Jinno, Y., Narita, H., Yonezawa, T., Nakagawa, A., & Okamura, Y. (2022) Interaction between S4 and the phosphatase domain mediates electrochemical coupling in voltage-sensing phosphatase (VSP). Proc. Natl. Acad. Sci. USA, 119, e2200364119.
34) Fernández-Mariño, A.I., Tan, X.F., Bae, C., Huffer, K., Jiang, J., & Swartz, K.J. (2023) Inactivation of the Kv2.1 channel through electromechanical coupling. Nature, 622, 410–417.
35) Yao, H., Mehta, V., Gulati, A., & Drew, D. (2023) Structure and electrochemical coupling of a voltage-gated Na+/H+ exchanger. Nature, 623, 193–201.
36) Kurokawa, T., Takasuga, S., Sakata, S., Yamaguchi, S., Horie, S., Homma, K.J., Sasaki, T., & Okamura, Y. (2012) 3′ Phosphatase activity toward phosphatidylinositol 3,4-bisphosphate [PI(3,4)P2] by voltage-sensing phosphatase (VSP). Proc. Natl. Acad. Sci. USA, 109, 10089–10094.
37) Grimm, S.S. & Isacoff, E.Y. (2016) Allosteric substrate switching in a voltage-sensing lipid phosphatase. Nat. Chem. Biol., 12, 261–267.
38) Zeng, X.H., Yang, C., Kim, S.T., Lingle, C.J., & Xia, X.M. (2011) Deletion of the Slo3 gene abolishes alkalizationactivated K+ current in mouse spermatozoa. Proc. Natl. Acad. Sci. USA, 108, 5879–5884.
39) Neuhaus, H. & Hollemann, T. (2009) Kidney specific expression of cTPTE during development of the chick embryo. Gene Expr. Patterns, 9, 568–571.
40) Ogasawara, M., Sasaki, M., Nakazawa, N., Nishino, A., & Okamura, Y. (2011) Gene expression profile of Ci-VSP in juveniles and adult blood cells of ascidian. Gene Expr. Patterns, 11, 233–238.
41) Park, J., Levic, D.S., Sumigray, K.D., Bagwell, J., Eroglu, O., Block, C.L., Eroglu, C., Barry, R., Lickwar, C.R., Rawls, J.F., et al. (2019) Lysomsome-Rich Enterocytes Mediate Protein Absorption in the Vertebrate Gut. Dev. Cell, 51, 7–20.e6.
42) Ratanayotha, A., Matsuda, M., Kimura, Y., Takenaga, F., Mizuno, T., Hossain, M.I., Higashijima, S.I., Kawai, T., Ogasawara, M., & Okamura, Y. (2022) Voltage-sensing phosphatase (Vsp) regulates endocytosis-dependent nutrient absorption in chordate enterocytes. Commun. Biol., 5, 948.
43) Yamamoto, T. (1966) An electron microscope study of the columnar epithelial cell in the intestine of fresh water teleosts: goldfish (Carassius auratus) and rainbow trout (Salmo irideus). Cell Tissue Res., 72, 66–87.
44) Siegel, M.S. & Isacoff, E.Y. (1997) A genetically encoded optical probe of membrane voltage. Neuron, 19, 735–741.
45) Tsutsui, H., Mizutani, N., & Okamura, Y. (2021) Engineering voltage sensing phosphatase (VSP). Methods Enzymol., 654, 85–114.
46) Okamura, Y. & Yoshioka, D. (2023) What voltage-sensing phosphatases can reveal about the mechanisms of ion channel regulation by phosphoinositides. Biochem. Soc. Trans., 51, 827–839.
47) Kawanabe, A., Mizutani, N., Polat, O.K., Yonezawa, T., Kawai, T., Mori, M.X., & Okamura, Y. (2020) Engineering an enhanced voltage-sensing phosphatase. J. Gen. Physiol., 152, e201912491.
48) Lacroix, J., Halaszovich, C.R., Schreiber, D.N., Leitner, M.G., Bezanilla, F., Oliver, D., & Villalba-Galea, C.A. (2011) Controlling the activity of a phosphatase and tensin homolog (PTEN) by membrane potential. J. Biol. Chem., 286, 17945–17953.
49) Mandala, V.S. & MacKinnon, R. (2023) The membrane electric field regulates the PIP2-binding site to gate the KCNQ1 channel. Proc. Natl. Acad. Sci. USA, 120, e2301985120.
50) Tsutsui, H., Jinno, Y., Mizutani, N., & Okamura, Y. (2024) Structural change of the cytoplasmic N-terminus and S1 segment of voltage-sensing phosphatase reported by Anap. Acta Physiol. (Oxf.), e14137.
51) Paixao, I.C., Mizutani, N., Matsuda, M., Andriani, R.T., Kawai, T., Nakagawa, A., Okochi, Y., & Okamura, Y. (2023) Role of K364 next to the active site cysteine in voltage-dependent phosphatase activity of Ci-VSP. Biophys. J., 122, 2267–2284.
52) Wei, Y., Stec, B., Redfield, A.G., Weerapana, E., & Roberts, M.F. (2015) Phospholipid-binding sites of phosphatase and tensin homolog (PTEN): Exploring the mechanism of phosphatidylinositol 4,5-bisphosphate activation. J. Biol. Chem., 290, 1592–1606.
53) Villalba-Galea, C.A., Miceli, F., Taglialatela, M., & Bezanilla, F. (2009) Coupling between the voltage-sensing and phosphatase domains of Ci-VSP. J. Gen. Physiol., 134, 5–14.
54) Kohout, S.C., Bell, S.C., Liu, L., Xu, Q., Minor, D.L. Jr., & Isacoff, E.Y. (2010) Electrochemical coupling in the voltage-dependent phosphatase Ci-VSP. Nat. Chem. Biol., 6, 369–375.
著者紹介Author Profile
 岡村 康司(おかむら やすし)
岡村 康司(おかむら やすし)大阪大学大学院医学系研究科 教授.医学博士.
略歴1960年東京都江戸川区に生る.85年東京大学医学部卒業.東京大学助手,産業技術総合研究所主任研究員,自然科学研究機構岡崎統合バイオサイエンスセンター教授を経て,2008年より現職.
研究テーマと抱負真っ暗の中で,透過型電子顕微鏡の蛍光板に映し出された神経細胞の精緻な構造に驚いたのと,刻一刻変化していく細胞の生の電気現象(ミリ秒から,日のオーダーまで)に感銘したのが研究生活の入り口.現在電位センサーをもつタンパク質やイオンチャネルについて,構造と機能,恒常性維持,生物機能を横断して研究しています.
ウェブサイトhttp://www.med.osaka-u.ac.jp/pub/phys2/okamura/
趣味散策,歴史,文学.