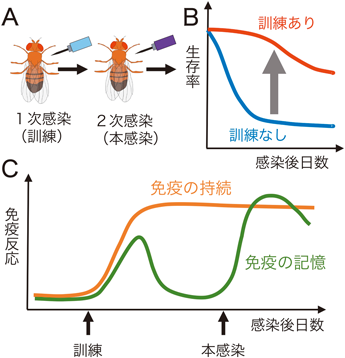
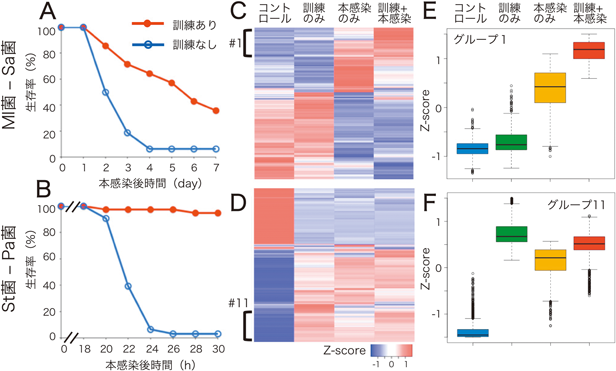
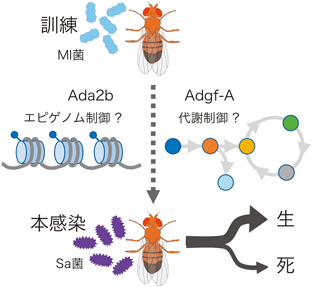
自然免疫の記憶における遺伝子発現制御Regulatory mechanism of gene expression in innate immune memory
東北大学大学院薬学研究科Graduate School of Pharmaceutical Sciences, Tohoku University ◇ 〒980–8578 仙台市青葉区荒巻字青葉6–3 ◇ 6–3 Aoba, Aramaki, Aoba-ku, Sendai 980–8578, Japan
発行日:2024年4月25日Published: April 25, 2024