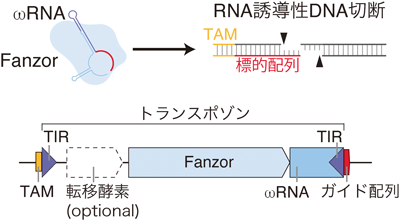
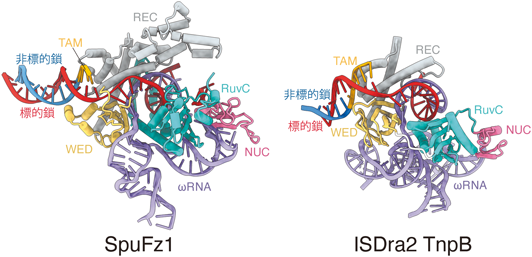
真核生物初のCRISPR-Cas様酵素の発見Discovery of the first CRISPR-Cas-like enzyme in eukaryotes
ブロード研究所,フェン・チャン研究室Broad Institute of MIT and Harvard, Feng Zhang Lab ◇ マサチューセッツ州ケンブリッジ,75エイムスストリート ◇ 75 Ames Street, Cambridge, MA 02142, U.S.
発行日:2024年4月25日Published: April 25, 2024